Interaktiivinen geneettinen maailmankartta. Venäjän ukrainalaisten, valkovenäläisten ja tataarien, slaavien ja valkoihoisten, juutalaisten, suomalaisten ja muiden populaatioiden genetiikka
Tässä tutkimuksessa käytetään yli 80 000 autosomaalista viipaletta, referenssipistettä - tämä on erittäin korkea resoluutio, jonka avulla voit havaita jopa suhteellisen pienet geneettiset vaikutukset suurimmassa osassa ihmisiä. Vertailevat analyysitiedot on otettu geneettisten komponenttien vertailevan analyysin asiantuntijan V. Verenichin avoimesta tutkimuksesta. Itse geneettiset laskimet sijaitsevat GedMatch-palvelussa, ja niiden avulla kuka tahansa voi saada selville oman vertailukohtansa geneettisestä kaaviosta. Tätä varten riittää, että sinulla on FTDNA:n tai 23andMe:n autosomaalisen testin tulokset. Tutkimuksen lopussa on MDLP World-22 -projektin tärkeimpien autosomaalisten komponenttien maantieteellisen jakauman ja taajuusmaksimien kartat.
Alla olevissa kaavioissa esitetään pääkomponentit ja niiden keskimääräinen prosenttiosuus kustakin populaatiosta. Yksi rivi on prosentuaalinen erittely yhdelle populaatiolle. Jokainen jako (pystypalkki) vastaa 10 %, ja autosomaalisten komponenttien nimet ovat samassa järjestyksessä vasemmalta oikealle kuin selitteessä ylhäältä alas. Mitä samanlaisempi prosenttiosuus kokonaisgenetiikkasta eri kansoilla on, sitä samanlaisemmalta näyttää annetussa kaaviossa oleva luku. Joten aloitetaan...
Saksalaisten, liettualaisten, venäläisten, ruotsalaisten, suomalaisten jne.
Tämä kaavio näyttää tärkeimmät geneettiset komponentit eurooppalaisille kansoille, ja se on kohdistettu Itä-Euroopan komponentin (Koillis-Euroopan) vähenemiseen eri populaatioissa. Kuten näette, kaikki Euroopan kansat ovat geneettisesti melko erilaisia, ja koska niiden geneettiset komponentit ovat samaa alkuperää, ne ovat kuitenkin hyvin eri prosenttiosuuksia. Kaikille slaaveille ja balttialaisille yleensä yksi merkittävimmistä on tämä Itä-Euroopan komponentti, joka on liettualaisilla ja valkovenäläisillä maksimissaan. Luultavasti arkeologisen "langallisen tavarakulttuurin" ajoista lähtien näiden maiden alue on ollut tämän komponentin alkuperäkeskus. Sitä edustaa yli 80 % liettualaisista ja vain 20 % italialaisista.Violetti edustaa Atlanto-Välimeren komponenttia, ja se lisääntyy, kun siirryt koillisesta lounaaseen. Suomalaisilla se on siis keskimäärin 15 % ja italialaisilla 40 %. Muut komponentit ovat vähemmän korostettuja.
Venäjän ukrainalaisten valkovenäläisten genetiikka

Tämä kaavio näyttää itäslaavit - venäläisiä, valkovenäläisiä, ukrainalaisia... Huomio kiinnitetään kolmen lueteltujen kansan geneettisten kuvioiden samankaltaisuuteen, ja virhemarginaalien sisällä ne eroavat melko merkityksettömästi - ukrainalaisilla ja etelävenäläisillä Länsi-Aasian komponentti on hieman lisääntynyt ja pohjoisvenäläisillä. on yhden Siperian komponentin, jota kutsutaan ehdollisesti samojediksi, hienoinen nousu ja Euroopan mesoliittisen komponenttien nousu noin 10 prosenttiin, mikä jälkimmäisen indikaattorin mukaan tuo ne lähemmäksi Skandinavian saksankielistä väestöä - ruotsalaiset.
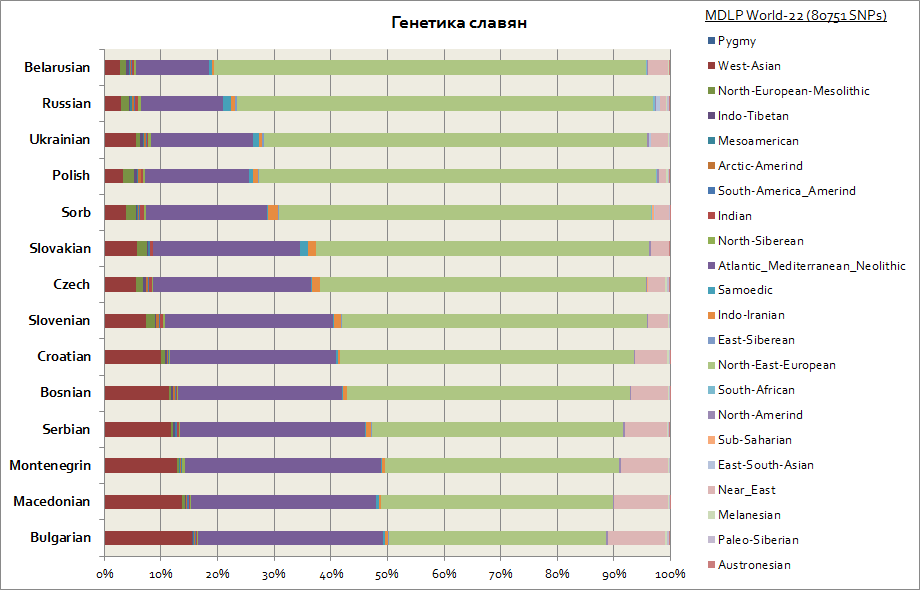
Tämä kaavio kuvaa kaikkia slaaveja, mukaan lukien läntiset - puolalaiset ja tšekit, sekä eteläiset - serbit, bulgarialaiset, makedonialaiset jne.
Kaikkien slaavien pääkomponentteja on 2. Nämä ovat Itä-Euroopan ja Atlantin ja Välimeren alueen. Ensimmäinen on korkeintaan valkovenäläisille ja toinen kaikille eteläslaaville - serbeille, makedonialaisille, bulgarialaisille. Itäeurooppalainen komponentti on alkuperältään ensisijaisempi slaavien keskuudessa, ja Atlanto-Välimeri on hankittu enemmän, kun slaavit muuttivat Balkanille. Länsi-ukrainalaisilla ja slovakeilla samojedikomponentin kasvu on heikkoa verrattuna naapurimaiden slaavikansoihin - valkovenäläisiin, tšekkeihin, puolalaisiin; se on luultavasti geneettinen jälki hunnien ja ugrilaisten keskiaikaisesta muuttoliikkeestä Keski-Eurooppaan.
Slaavien, venäläisten ja tataarien, saksalaisten, valkoihoisten, juutalaisten jne.
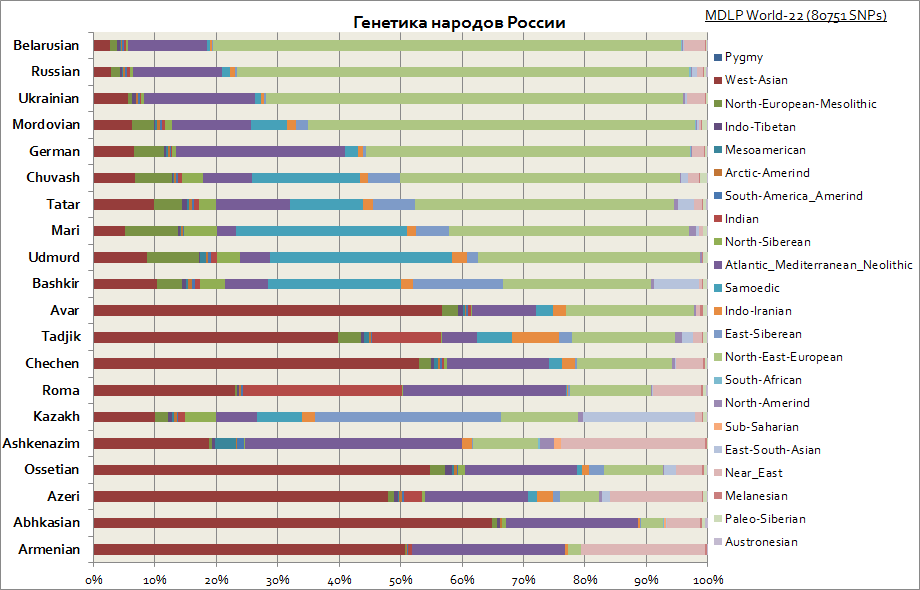
Tämä kaavio näyttää Venäjän kansojen erilaiset alkuperät. Kuten näette, slaavien keskuudessa Itä-Euroopan komponentti on tärkein, ja Volgan alueen kansojen keskuudessa Siperian komponenttien osuus kasvaa. Kaukasialaisille tyypillisin on Länsi-Aasialainen komponentti, Välimeri ja Lähi-itä.
Suomalaisten, ugrilaisten, udmurtien, unkarilaisten, saamelaisten jne.

Kuten näette, suomalaisille, vepsalaisille ja karjalaisille on ominaista samanlainen geneettinen alkuperä kuin slaaveilla. Niissä on myös korkein itäeurooppalainen komponentti, joka laskee lähemmäs Uralia ja Volgan aluetta, ja Siperian komponentit lisääntyvät tällä alueella. Kaikilla suomalais-ugrilaisilla kansoilla on myös merkittävä osa Euroopan mesoliittia, joka saavuttaa lähes 80 % saamelaisista ja liittyy Euroopan esi-indoeurooppalaiseen ja esineoliittiseen väestöön. Unkarilaisille kokonaisuudessaan on ominaista sarja samoja geneettisiä komponentteja kuin muille Karpaattien alueen ja Keski-Euroopan populaatioille.
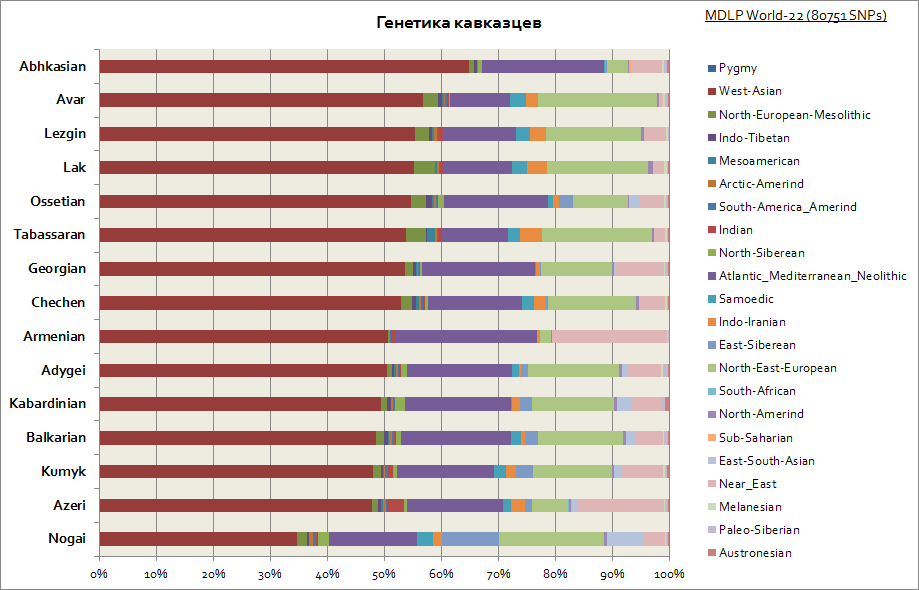
Kuten näette, koko Kaukasukselle on ominaista suhteellisen samanlainen geneettinen alkuperä - tämä on suuri osuus Länsi-Aasian komponentista ja Välimerestä. Vain Nogait ovat hieman tyrmistyneet - niiden osuus Siperian komponenteista kasvaa.

Kuten voidaan nähdä, aškenatsimilla ja sefardeilla on suuri taajuus Länsi-Aasian, Atlantin ja Välimeren alueen ja Lähi-idän komponentteja. Samaan aikaan aškenasien Siperian komponentti on hieman lisääntynyt, mikä todennäköisesti liittyy Khazar-perintöön ja jopa 30 prosentin kasvuun Itä-Euroopan komponentista, mikä tämän indikaattorin mukaan tuo heidät lähemmäksi Etelä-Euroopan maissa.
Varsinkin heidän "yrityksestään" vain Etiopian juutalaiset ja Intian juutalaiset. Ensin mainituissa on suuri osuus Saharan eteläpuolisesta Afrikasta (jopa 40 %), kun taas jälkimmäisissä on suuri osuus eteläaasialaista geneettistä komponenttia, jota kutsutaan perinteisesti intialaiseksi (jopa 50 %).
Tataarien, baškiirien, azerbaidžanilaisten, tšuvashien jne. genetiikka.
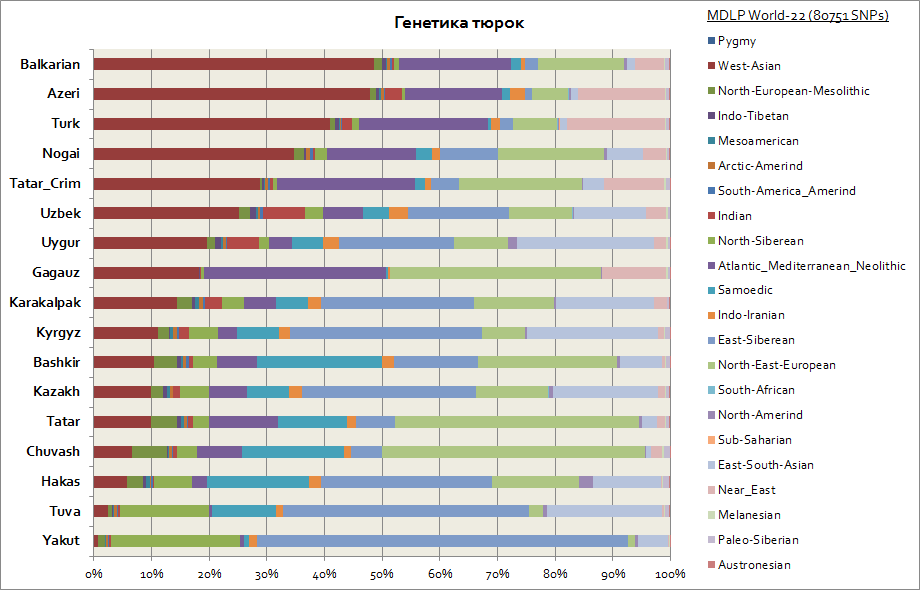
Geneettisesti turkkilaiset osoittautuivat yhdeksi heterogeenisimmista etnisistä ryhmistä, joten niiden geneettiset komponentit eroavat toisistaan merkittävästi. Joten, koska turkkilaisten ensisijainen kotimaa on Siperia, jakutit, tuvinalaiset ja hakassit säilyttivät suurimman osan Itä-Siperian autosomaalisesta komponentista, joka on 30–65%. Tämä geneettinen komponentti on myös tärkein kirgissien ja kazakstien keskuudessa. Loput komponentit tuovat turkkilaisia lähemmäksi asuinalueiden kansoja. Jakutit ja tuvanit ovat siis Pohjois-Siperian ja Samodian komponentteja. Yhteensä nämä 3 siperialaista komponenttia jakuuteilla jopa 90 %, tuvinalaisilla jopa 70 %, ja Itä-Etelä-Aasian komponentti kasvaa jopa 20 %, mikä liittyy enemmän idän väestön muuttovirtoihin. Aasia. Baškiirien kohdalla kolmen siperialaisen komponentin osuus on jopa 45 % ja Kaakkois-Aasian komponentin osuus jopa 10 %. Tataareilla on tietoja kolmesta siperialaisesta geneettisestä komponentista keskimäärin 25-50%. Samanaikaisesti valkoihoiselle väestölle ominaisten komponenttien osuus baškiirien joukossa on jopa 45%, ja tataarien keskuudessa keskimäärin 50 - 70%. Azerbaidžanilaisten ja turkkilaisten genetiikka ei käytännössä eroa virhemarginaalin sisällä, heillä, kuten muilla Kaukasuksen ja Transkaukasian alueen kansoilla, on merkittävä Länsi-Aasialainen komponentti (jopa 50 %) ja Atlantin ja Välimeren alue. keskimäärin jopa 20 %). Azerbaidžanilaiset, turkkilaiset ja balkarit esittävät kolmen siperialaisen komponentin osuuden - tasolla 3-7%.
Johtopäätös
Kansojen genetiikalla ei ole suoraa ja merkittävää korrelaatiota kieliperheiden jakautumisen tai homogeenisten markkerien - Y-DNA- ja mt-DNA-haploryhmien - prosenttiosuuden kanssa, jotka ovat edustettuina tietyssä populaatiossa. Suurin korrelaatio voidaan jäljittää aluemaantieteellisen periaatteen mukaisesti. Siperialaisten komponenttien osuus mongoloidirodulle kokonaisuudessaan siis pienenee vähitellen idästä länteen ja kaukasialaiselle rodulle ominaisten komponenttien osuus kasvaa vastaavasti. Uralin pohjoisosasta Keski-Aasiaan kulkevan linjan raja-alueilla niiden suhde on suunnilleen sama. Baikal-järven itäpuolella olevilla alueilla suurelle kaukasialaiselle rodulle ominaisia geneettisiä komponentteja ei käytännössä ole edustettuina, samaan aikaan Pechora-Volgan alueen länsipuolisilla alueilla suurelle mongoloidirodulle ominaiset siperialaiset komponentit katoavat.Itäeurooppalaisen geneettisen komponentin leviäminen Siperiaan tapahtui suurelta osin jo pronssikaudella (Andronov-ympyrän kulttuuri), vaikka yksittäiset huiput Siperian äärimmäisessä itäosassa tšuktšien keskuudessa saattoivat liittyä venäläisten muuttoihin vuonna 1700-luvulla.
Negroidirodulle ominaisen Saharan eteläpuolisen komponentin osuus on jakautunut koko Afrikkaan - eteläiselle Välimerelle ja Afrikan mantereen pohjoisrajalle asti saavuttaen maksiminsa päiväntasaajan osassa, eikä sitä käytännössä koskaan esiinny sen ulkopuolella; vaalea tausta yleinen Arabian niemimaalla ja Iranin tasangon eteläosassa.
Geneettisten komponenttien maantiede
Aleksei Zorrin
Projekti
Geenianalyysi on jo pitkään ja lujasti tullut rutiinitoimenpiteiden joukkoon sellaisilla soveltavilla tiedon aloilla kuin oikeuslääketiede ja lääketiede. Geneettisten markkerien tunnistusmenetelmien parantaminen löytää pääsääntöisesti käyttöä tieteellisissä perusongelmissa.
Uusien teknisten ratkaisujen ansiosta mm. täysin sekvensoitu mtDNA esihistoriallinen neandertalilainen ihminen, ja lähitulevaisuudessa tutkijat toivovat voivansa tulkita sen täydellisen ydingenomia ja verrata sitä ihmiseen. Oma historiamme paljastuu meille yhä enemmän ei arkeologisten löytöjen, vaan geneettisen analyysin menetelmien kautta. Opimme esimerkiksi Maan asettumisen likimääräisen kronologian ja selvisimme, että ihmiskunta oli sen kehityksen varhaisessa vaiheessa melkein katosi maan pinnalta kuivuuden vuoksi.
Useisiin miljardeihin DNA:n nukleotideihin koodattu data voi kuitenkin kertoa paljon enemmän tietystä henkilöstä ja koko ihmiskunnasta. John Novembre Kalifornian yliopistosta Los Angelesista ja hänen kollegansa osoittivat, kuinka yli kolmen tuhannen eurooppalaisen laajamittainen geneettinen analyysi on jo mahdollistanut Euroopan väestörakenteen paljastamisen ja kuinka geneettisesti erilaisia pienten eurooppalaisten edustajia. osavaltiot ovat.
Nyt tiedemiehet voivat useiden satojen kilometrien tarkkuudella määrittää minkä tahansa eurooppalaisen alkuperäpaikan vain hänen DNA:nsa perusteella.
On olemassa monia matemaattisia menetelmiä tämän epäselvästi muotoillun ongelman ratkaisemiseksi; genetiikassa yleisin on pääkomponenttimenetelmä, jonka avulla moniulotteisessa tietojoukossa voidaan määrittää sellaiset "akselit", joilla genomierot näkyvät eniten. Novembre ja hänen kollegansa käyttivät sitä hyväkseen ja rajoittivat itsensä kahteen tällaiseen akseliin.
Sijoittaessaan kaikki lähes viisitoistasataa pistettä koneeseen he näkivät kuvan, joka muistutti hätkähdyttävästi Euroopan poliittista karttaa.
Portugalilaiset ja espanjalaiset genomit olivat tällä kartalla ranskalaisten vasemmalla puolella, saksalaiset - oikealla, "italialaiset" sijaitsivat ranskalaisten alla ja britit heidän yläpuolellaan. Onneksi tuloksena olevaa karttaa voi kiertää haluamallaan tavalla - tämä ei muuta pisteiden sijaintien välisiä geometrisia suhteita. Joten tutkijoiden täytyi vain verrata geneettistä kuvaajaa maantieteelliseen karttaan - niin sanottua kliinisuutta pääakselia vastaavan yhdistelmäparametrin mukaan - ja määrittää, että tärkein "pääkomponentti" vastaa etelä-kaakkoon - pohjois-luoteis-akselilla, ja nimittäin tässä muodossa ja esittele kartta kollegoille.
John Novembren ja hänen kollegoidensa Euroopan geneettinen kartta. Alhaalla oikealla - vähemmän silmiinpistävä tulos Manfred Kaiserin samankaltaisesta työstä, joka julkaistiin kuukausi sitten. // John Novembre / M.Kaiser
Lopullinen kartta ei tietenkään ollut ilman puutteita. Paitsi, että yksittäiset genomit ovat levinneet kauas niitä vastaavien maiden rajojen ulkopuolelle. Venäläiset asettuivat tälle kaaviolle tšekkien ja puolalaisten keskuudessa ja löysivät itsensä paljon ukrainalaisten "länteen" puolelle, kun taas slovakit putosivat yleensä syvälle Alppien alle ja joutuivat aivan italialaisen "saappaan" päähän. Tämä selittyy kuitenkin molempien niukan otoksen erityispiirteillä: venäläisiä oli alkutiedoissa vain kuusi ja slovakialaisia vain yksi.
Siitä huolimatta, käyttämällä vain tätä karttaa, 90% ihmisistä kotimaa voidaan osoittaa 700 km:n tarkkuudella ja 50% - jopa 300 km:n tarkkuudella.
Tämä huolimatta siitä, että 200 tuhatta "kirjainta" DNA:ta - alle 0,01% koko ihmisen geneettisestä koodista.
Kaaviosta näkyy myös hyvin selvästi, että eurooppalaisten-eteläisten geneettinen monimuotoisuus on paljon suurempi kuin pohjoisten kansojen. Tämä data, Novembran itsensä mielestä, voidaan helposti selittää ihmisten Euroopan asuttamisen historialla. Sen jälkeen kun Homo sapiens ilmestyi tänne 35 tuhatta vuotta sitten, vain 15 tuhatta vuotta myöhemmin, jäätiköiden vetäytyessä, tapahtui ensimmäinen kansojen massamuutto pohjoiseen, ja vielä 10 tuhannen vuoden kuluttua maatalouden syntyessä uusia eteläisten ryhmät menivät pohjoiseen, jotka kaikki nämä vuosituhannet edustivat vaikuttavia, hyvin muodostuneita populaatioita.
Muuten samanlainen Työ, joka perustuu suureen osaan samoihin geneettisiin tietoihin, julkaistiin Current Biology -lehdessä kuukausi sitten. On syytä huomata, että sen johtaja Manfred Kaiser onnistui saamaan samanlaisen, vaikkakaan ei niin mehukkaan kuvan käyttämällä samaa pääkomponenttien menetelmää. Analyysi sisälsi 2 500 genomia, jotka GlaxoSmithKline, geenien metsästysyritys, joka vastaa ihmisten huumeiden käytön sivuvaikutuksista, kokosi huolellisesti.

Sveitsin väestön "geneettinen kartta" // John Novembre
Samanaikaisesti pääkomponenttien menetelmä ei kummassakaan tapauksessa kyennyt jakamaan eurooppalaisia erillisiin geneettisiin klustereihin. Toisin sanoen eurooppalaisten geneettinen monimuotoisuus on niin pieni, että edes puoli miljoonaa mononukleotidipolymorfismia ei riitä jakamaan vain kolmetuhatta ihmistä pieniin alaryhmiin.
"Puhumme joskus mielellämme" venäläisistä geeneistä "," puolalaisista geeneistä "tai" tataarin "geeneistä. Mutta tämä artikkeli osoittaa meille selvästi, että sellaisia geenejä ei ole!"
- kommentoi näitä töitä Gazeta.Ru:lle, joka on yksi tärkeimmistä kansallisista genogeografian asiantuntijoista Oleg Balanovsky Venäjän lääketieteellisen akatemian lääketieteellisen genetiikan tutkimuskeskuksen ihmispopulaatiogenetiikan laboratoriosta, hyvin tunnettu lukijoillemme.
"Vasta satatuhatta geeniä tutkittuaan kirjoittajat pystyivät (ja sitten useiden satojen kilometrien virheellä) määrittämään tutkittujen eurooppalaisten todennäköiset alkuperäpaikat. Tämä on mahdotonta yhdellä geenillä tai jopa sadalla geenillä! - sanoo Balanovsky. - Tietyn maan väestölle ei ole ominaista yhtä ainoaa geeniä. Vain satojen tuhansien geenien yhdistelmä on enemmän tai vähemmän spesifinen.
Tämä tarkoittaa, että päättely niille, jotka pelkäävät vaikutus ihmisten geenipooliin erityisten geeniensä kautta."
"Olemme odottaneet tällaista artikkelia pitkään. Miljoonan geneettisen markkerin analysointitekniikka on ollut olemassa useita vuosia. Ja tässä artikkelissa näitä uusia tekniikoita sovelletaan vihdoin Euroopan geenipoolin tutkimiseen, tutkija jatkaa. - Tämä työ osoittaa täydellisesti tieteen jatkuvuuden: kunhan 10-15 vuotta sitten tehtiin töitä mitokondrio-DNA:sta (äidin perinnöllisyyslinja) ja Y-kromosomista (isän perintölinja), ensimmäinen läpimurto tapahtui myös geenissä. Euroopan joukossa, ja jopa päätelmä maantieteen tärkeydestä geenipoolin järjestykseen tehtiin samalla tavalla kuin nyt."
Balanovskin mukaan nykytekniikat mahdollistavat ennennäkemättömän luotettavuuden ja tulosten tarkkuuden: ”Mitä genetiikka tuskin pystyi erottamaan tiheässä sumussa neljäkymmentä vuotta sitten, jota kymmenen vuotta sitten alkoi valaista kaksi voimakasta sumuvaloa (mitokondrio-DNA ja Y-kromosomi). ), alamme nyt nähdä saman asian miljoonan heikomman valon valossa (geenejä, vaikkakin vähemmän informatiivisia, on lukuisia ja ne ovat jakautuneet koko genomiin).
"Seuraava ja viimeinen askel - täysi, sataprosenttinen valaistus - voidaan odottaa muutaman vuoden kuluttua, kun koko genomin täydellinen sekvensointi tulee saataville - ei miljoona nukleotidia, kuten nyt on, vaan kaikki genomissa olevat miljardit, venäläinen asiantuntija ennustaa.
Ainoa kysymys, joka hänen mielestään herää, on, onko tarpeen odottaa näin yksityiskohtaisia tietoja. Todellakin, jo nyt "kaksi voimakasta valoamme (mitokondrio-DNA ja Y-kromosomi) ja miljoona heikompaa valoa (mutta jakautuvat koko genomiin) antavat meille mahdollisuuden nähdä ihmisen geenipoolin rakenteen ja tehdä sekä perustavanlaatuisia tieteellisiä että sovellettavia johtopäätöksiä" .
Tiedemies on huolestunut siitä, että toistaiseksi tutkittujen geenien määrä genomissa on paljon suurempi kuin niiden ihmisten määrä, joilla näitä geenejä on tutkittu.
"Vain geenipoolin - koko geenijoukon - tutkiminen mahdollistaa ihmisten genetiikan tutkimisen auttaaksemme lääkäreitä tekemään diagnooseja ja kaikkia ihmisiä - jäljittämään oman kansansa tai oman perheensä historiaa." totesi Oleg Balanovsky.
5. tammikuuta 2013 Tulosta
Kuulemme jatkuvasti, että venäläiset eivät ole veren hitsaama kansa, veren sukulaiskunta, vaan joukko ihmisiä, joita yhdistää yhteinen kulttuuri ja alue. Kaikki muistavat Putinin iskulauseet "Ei ole puhtaita venäläisiä!" ja "raavi jokainen venäläinen, löydät varmasti tataarin."
Sanotaan, että olemme "verisesti hyvin erilaisia", "emme itäneet yhdestä juuresta", vaan olimme sulatusuuni tataareille, kaukasialaisille, saksalaisille, suomalaisille, burjaateille, mordvaalaisille ja muille kansoille, jotka koskaan tulivat, tulivat, saapuivat maamme, ja otimme ne kaikki vastaan, päästimme ne taloon, veimme sukulaisillemme.
Tästä on tullut lähes aksiooma venäjän käsitettä laimentavien poliitikkojen keskuudessa, ja samalla kaikille siitä on tullut pääsylippu Venäjän kansan ympäristöön.
Tämä lähestymistapa, jonka monet russofobiset à la "ihmisoikeusjärjestöt" ja venäläiset russofobiset tiedotusvälineet ovat nostaneet lippuun, on tulvinut ilmassa. Mutta Putinin ja muiden hänen kaltaistensa on ennemmin tai myöhemmin silti vastattava Venäjän kansaa kohtaan osoittamistaan nöyryyttävistä sanoistaan. Tiedemiesten tuomio on armoton:
1) Vuonna 2009 venäläisen etnoksen edustajan genomin täydellinen "luku" (sekvensointi) valmistui. Eli kaikkien kuuden miljardin nukleotidin sekvenssi venäläisen miehen genomissa on määritetty. Hänen geenitaloutensa on nyt täysin näkyvissä.
(Ihmisen genomi koostuu 23 kromosomiparista: 23 - äidiltä, 23 - isältä. Jokainen kromosomi sisältää yhden DNA-molekyylin, joka muodostuu 50-250 miljoonan nukleotidin ketjusta. Venäläisen miehen genomi sekvensoitiin. center "Kurchatov-instituutti", Venäjän tiedeakatemian kirjeenvaihtajajäsenen, kansallisen tutkimuskeskuksen "Kurchatov-instituutin" johtajan Mihail Kovaltšukin aloitteesta Venäjän tiedeakatemiasta saatujen tietojen mukaan vain sekvensointilaitteiden hankintaa varten, Kurchatov-instituutti käytti noin 20 miljoonaa dollaria. Keskus "Kurchatov-instituutti" on tunnustettu tieteellinen asema maailmassa.)
Tiedetään, että tämä on seitsemäs dekoodattu genomi Uralin harjanteen takana: ennen sitä oli jakuteja, burjaatteja, kiinalaisia, kazakseja, vanhauskoisia, hanteja. Eli kaikki edellytykset Venäjän ensimmäiselle etniselle kartalle on luotu. Mutta kaikki nämä olivat niin sanotusti yhdistelmägenomeja: kappaleita, jotka koottiin dekoodattuaan saman populaation eri edustajien geneettisen materiaalin.
Tietyn venäläisen miehen täydellinen geneettinen muotokuva on vasta kahdeksas maailmassa. Nyt on joku, johon verrata venäläisiä: amerikkalainen, afrikkalainen, korealainen, eurooppalainen ...
« Emme löytäneet havaittavia tataarien panoksia venäjän genomissa, mikä kumoaa teoriat mongolien ikeen tuhoisasta vaikutuksesta, - korostaa Kansallisen tutkimuskeskuksen "Kurchatov-instituutin" genomisen suunnan johtaja, akateemikko Konstantin Skryabin. -Siperialaiset ovat geneettisesti identtisiä vanhauskoisten kanssa, heillä on yksi venäläinen genomi. Venäläisten ja ukrainalaisten genomien välillä ei ole eroja - yksi genomi... Eromme puolalaisten kanssa ovat vähäiset."
Akateemikko Konstantin Skryabin uskoo, että "geneettinen kartta kaikista maailman kansoista kootaan viidessä-kuudessa vuodessa - tämä on ratkaiseva askel kohti minkä tahansa etnisen ryhmän herkkyyttä lääkkeille, sairauksille ja elintarvikkeille." Tunne mitä se maksaa ... Amerikkalaiset 1990-luvulla antoivat seuraavat arviot: yhden nukleotidin sekvensoinnin kustannukset - 1 dollari; muiden lähteiden mukaan - jopa 3-5 dollaria.
(Mitokondrion DNA:n ja ihmisen Y-kromosomin DNA:n sekvensointi (geneettisen koodin täsmentäminen) on tähän mennessä edistynein DNA-analyysimenetelmä. välittyy myös käytännössä muuttumattomana urospuolisille jälkeläisille, kun taas kaikki muut kromosomit, jotka siirtyvät isältä ja äidiltä lapsilleen, sekoitetaan luonnostaan korttipakan tavoin ennen jakamista. , toisin kuin epäsuorat merkit (ulkonäkö, kehon mittasuhteet), sekvensointi mitokondrio-DNA ja Y-kromosomin DNA on kiistaton ja suora todiste ihmisten sukulaisuusasteesta.)
2) Erinomainen antropologi, ihmisen biologisen luonnon tutkija, A.P. Bogdanov kirjoitti 1800-luvun lopulla: "Käytämme usein ilmaisuja: tämä on puhtaasti venäläistä kauneutta, tämä on sylkevä jänis, tyypillisesti venäläiset kasvot. Voidaan vakuuttua siitä, ettei tässä venäläisen fysiognomian yleisessä ilmaisussa ole mitään fantastista, vaan todellista. Jokaisessa meistä "tietoisuuden" alueella on melko selvä käsite venäläistyypistä "(AP Bogdanov" Antropologinen fysiognomia ". M., 1878).
Sata vuotta myöhemmin ja nyt moderni antropologi V. Deryabin tulee uusimman sekaominaisuuksien matemaattisen monimuuttuja-analyysin avulla samaan johtopäätökseen: "Ensimmäinen ja tärkein johtopäätös on todeta venäläisten merkittävä yhtenäisyys. koko Venäjällä ja mahdottomuus tunnistaa edes vastaavia alueellisia tyyppejä, jotka on selvästi erotettu toisistaan "(" Antropologian kysymyksiä ". Numero 88, 1995). Miten tämä venäläinen antropologinen yhtenäisyys, perinnöllisten geneettisten ominaisuuksien yhtenäisyys ilmaistaan ihmisen ulkonäössä, hänen ruumiinsa rakenteessa?
Ensinnäkin - hiusten väri ja silmien väri, kallon rakenteen muoto. Näiden merkkien mukaan me, venäläiset, eroamme sekä eurooppalaisista kansoista että mongoloideista. Eikä meitä voi verrata neekereihin ja seemiläisiin ollenkaan, erot ovat liian silmiinpistäviä. Akateemikko V.P. Alekseev osoitti suuren samankaltaisuuden kallon rakenteessa kaikkien nykyaikaisen venäläisen kansan edustajien kesken ja täsmensi samalla, että "protoslaavilainen tyyppi" on erittäin vakaa ja juontaa juurensa neoliittiseen aikakauteen ja mahdollisesti mesoliittiseen aikaan. . Antropologi Deryabinin laskelmien mukaan vaaleita silmiä (harmaa, harmaa-sininen, sininen ja sininen) löytyy venäläisistä 45 prosentista, Länsi-Euroopassa vaaleista vain 35 prosentista. Tummat, mustat hiukset venäläisillä on viidellä prosentilla, ulkomaisen Euroopan väestöstä 45 prosentilla. Perinteinen viisaus venäläisten "nenäkärkyydestä" ei myöskään ole vahvistunut. 75 prosentilla venäläisistä on suora nenäprofiili.
Antropologisten tutkijoiden johtopäätös:
”Venäläiset ovat rodultaan tyypillisiä valkoihoisia, joilla on keskeinen asema Euroopan kansojen joukossa useimpien antropologisten ominaisuuksiensa vuoksi ja eroavat hieman vaaleammasta silmien ja hiusten pigmentaatiosta. On myös välttämätöntä tunnustaa venäläisten rodullisen tyypin merkittävä yhtenäisyys koko Euroopan Venäjällä."
”Venäläinen on eurooppalainen, mutta eurooppalainen, jolla on vain hänelle ominaisia fyysisiä ominaisuuksia. Nämä merkit muodostavat sen, mitä kutsumme tyypilliseksi jäniksi."
Antropologit ovat vakavasti raapineet venäläistä, ja - venäläisissä ei ole tataaria, toisin sanoen mongoloidia. Yksi tyypillisistä mongoloidin merkeistä on epicanthus - mongolilainen poimu silmän sisäkulmassa. Tyypillisissä mongoloideissa tämä laskos löytyy 95 prosentista; kahdeksan ja puolen tuhannen venäläisen tutkimuksessa tällainen laskos löydettiin vain 12 ihmiseltä ja alkiomuodossa.
Toinen esimerkki. Venäläisillä on kirjaimellisesti erityinen veri - 1. ja 2. ryhmän ylivalta, mikä on osoituksena verensiirtoasemien pitkäaikaisesta käytännöstä. Esimerkiksi juutalaisilla vallitseva veriryhmä on 4., negatiivinen Rh-tekijä on yleisempi. Biokemiallisissa verikokeissa kävi ilmi, että venäläisillä, kuten kaikilla eurooppalaisilla kansoilla, on erityinen PH-c-geeni, tämä geeni käytännössä puuttuu mongoloideista (OV Borisova "Erytrosyyttihapon fosfataasin polymorfismi Neuvostoliiton eri väestöryhmissä". "Antropologian kysymykset". Numero 53, 1976).
Osoittautuu, että vaikka kuinka raapaisit venäläistä, ei silti tataaria, et löydä hänestä ketään muuta. Tämän vahvistaa tietosanakirja "People of Russia", luvussa "Venäjän väestön roturakenne" todetaan: "Kaukasialaisen rodun edustajat muodostavat yli 90 prosenttia maan väestöstä ja vielä 9 prosenttia. ovat valkoihoisten ja mongoloidien välillä sekoitettujen muotojen edustajia. Puhtaiden mongoloidien määrä ei ylitä 1 miljoonaa ihmistä. ("Peopeles of Russia". M., 1994).
On helppo laskea, että jos Venäjällä on 84 prosenttia venäläisiä, he ovat kaikki yksinomaan eurooppalaisia. Siperian, Volgan alueen, Kaukasuksen ja Uralin kansat ovat sekoitus eurooppalaista ja mongolialaista rotua. Tämän ilmaisi täydellisesti antropologi A.P. Bogdanov 1800-luvulla Venäjän kansoja tutkiessaan hän kirjoitti kumoamalla kaukaa, kaukaa tämän päivän myytin siitä, että venäläiset vuodattivat jonkun muun verta kansaansa hyökkäysten ja kolonisaation aikana:
”Ehkä monet venäläiset menivät naimisiin alkuperäiskansojen kanssa ja asettuivat asettumaan, mutta suurin osa primitiivisistä venäläisistä kolonisaattoreista kaikkialla Venäjällä ja Siperiassa ei ollut sellaisia. He olivat kauppiaita, teollisuuskansaa, jotka huolehtivat järjestäytymisestä omalla tavallaan oman vaurausihanteensa mukaisesti. Ja tämä venäläisen miehen ihanne ei ole ollenkaan sellainen, että se helposti vääristää hänen elämäänsä jollain "roskalla", koska nyt venäläinen mies kunnioittaa aivan liian usein pakanaa. Hän tekee bisnestä hänen kanssaan, hän on hellä ja ystävällinen hänen kanssaan, hän ystävystyy hänen kanssaan kaikessa, paitsi sukulaisuussuhteessa tuodakseen vieraan elementin perheeseensä. Tähän tavalliset venäläiset ovat edelleen vahvoja, ja mitä tulee perheeseen, kodin perustamiseen, täällä hänellä on eräänlainen aristokratia. Usein eri heimojen uudisasukkaat asuvat naapurustossa, mutta avioliitot heidän välillään ovat harvinaisia.
Vuosituhansien ajan venäläinen fyysinen tyyppi on pysynyt vakaana ja muuttumattomana, eikä se ole koskaan ollut maallamme toisinaan asuneiden heimojen risteymä. Myytti on hajonnut, meidän on ymmärrettävä, että veren kutsu ei ole tyhjä lause, että kansallinen käsityksemme venäläistyypistä on venäläisen rodun todellisuus. Meidän on opittava näkemään tämä rotu, ihailemaan sitä, arvostamaan sitä läheisten ja kaukaisten venäläisten sukulaisten keskuudessa. Ja sitten kenties venäläinen vetoomuksemme täysin vieraisiin ihmisiin, mutta omat kansamme meille - isä, äiti, veli, sisko, poika ja tytär - herää henkiin. Loppujen lopuksi olemme kaikki yhdestä juurista, yhdestä lajista - venäläisestä.
3) Antropologit pystyivät tunnistamaan tyypillisen venäläisen ihmisen ulkonäön. Tätä varten heidän oli käännettävä yhdeksi mittakaavaksi kaikki Antropologian museon valokuvakirjaston valokuvat maan venäläisten alueiden väestön tyypillisten edustajien koko kasvo- ja profiilikuvilla sekä yhdistettävä ne silmien pupillit päällekkäin. Lopulliset valokuvat osoittautuivat tietysti epäselviksi, mutta antoivat käsityksen referenssivenäläisten ulkonäöstä. Tämä oli ensimmäinen todella sensaatiomainen löytö. Loppujen lopuksi ranskalaisten tutkijoiden samanlaiset yritykset johtivat tulokseen, jonka heidän oli piilotettava maansa kansalaisilta: tuhansien yhdistelmien jälkeen saatujen viite Jacquesista ja Mariannesta saatujen valokuvien jälkeen he katselivat harmaita kasvottomia kasvojen soikeita. Sellainen kuva, jopa antropologiasta kaukaisimpien ranskalaisten keskuudessa, voisi herättää tarpeettoman kysymyksen: onko ranskalaista kansakuntaa ollenkaan?
Valitettavasti antropologit eivät menneet pidemmälle kuin loivat valokuvallisia muotokuvia maan eri alueiden venäläisen väestön tyypillisistä edustajista eivätkä asettaneet niitä toistensa päälle saadakseen absoluuttisen venäläisen miehen ulkonäön. Lopulta he joutuivat myöntämään, että he saattoivat joutua vaikeuksiin töissä tällaisesta valokuvasta. Muuten, Venäjän kansan "alueelliset" luonnokset julkaistiin yleisessä lehdistössä vasta vuonna 2002, ja ennen sitä ne julkaistiin pieninä painoina vain asiantuntijoille tarkoitetuissa tieteellisissä julkaisuissa. Nyt voit itse arvioida, kuinka samanlaisia ne ovat tyypillisten elokuvamaisten Ivanushkan ja Maryan kanssa.
Valitettavasti enimmäkseen mustavalkoiset vanhat arkistokuvat venäläisten ihmisten kasvoista eivät pysty välittämään venäläisen pituutta, ruumiinrakennetta, ihon, hiusten ja silmien väriä. Antropologit ovat kuitenkin luoneet verbaalisen muotokuvan venäläisistä miehistä ja naisista. He ovat keskivartaloisia ja keskipitkät, vaaleanruskeat hiukset ja vaaleat silmät - harmaat tai siniset. Muuten, tutkimuksen aikana saatiin myös suullinen muotokuva tyypillisestä ukrainalaisesta. Tavallinen ukrainalainen eroaa venäläisestä vain ihon, hiusten ja silmien värin osalta - hän on tummaihoinen brunette, jolla on säännölliset kasvonpiirteet ja ruskeat silmät. Nuhakas nenä osoittautui täysin epätyypilliseksi itäslaavilaisille (löytyy vain 7%:lla venäläisistä ja ukrainalaisista), tämä merkki on tyypillisempi saksalaisille (25%).
4) Venäjän perustutkimussäätiö myönsi vuonna 2000 valtion budjettivaroista noin puoli miljoonaa ruplaa Venäjän kansan geenipoolin tutkimukseen. On mahdotonta toteuttaa vakavaa ohjelmaa sellaisella rahoituksella. Mutta se oli enemmän maamerkki kuin pelkkä taloudellinen päätös, mikä osoitti muutosta maan tieteellisissä prioriteeteissa. Ensimmäistä kertaa Venäjän historiassa Venäjän lääketieteen akatemian lääketieteellisen genetiikan keskuksen ihmispopulaatiogenetiikan laboratorion tutkijat, jotka saivat apurahan Venäjän perustutkimussäätiöltä, pystyivät keskittymään täysin tutkimukseen Venäjän kansan, ei pienten kansojen, geenipooli kolmen vuoden ajan. Ja rajallinen rahoitus vain vauhditti heidän kekseliäisyyttään. He täydensivät molekyyligeneettisiä tutkimuksiaan analyysillä venäläisten sukunimien esiintymistiheydestä maassa. Tämä menetelmä oli erittäin halpa, mutta sen tietosisältö ylitti kaikki odotukset: sukunimien maantieteen vertailu geneettisten DNA-merkkien maantieteeseen osoitti niiden lähes täydellisen yhteensopivuuden.
Valitettavasti tiedotusvälineissä ilmestyneet tulkinnat perheanalyysistä sen jälkeen, kun tiedot oli julkaistu ensimmäisen kerran erikoistuneessa tieteellisessä lehdessä, saattoivat luoda väärän kuvan tutkijoiden valtavan työn tavoitteista ja tuloksista. Projektin johtaja, tieteiden tohtori Elena Balanovskaja selitti, että pääasia ei ollut se, että sukunimi Smirnov osoittautui yleisemmaksi venäläisten keskuudessa kuin Ivanov, vaan että ensimmäistä kertaa täydellinen luettelo aidosti venäläisistä sukunimistä oli koottu maan alueille. Ensin laadittiin luettelot viidelle ehdolliselle alueelle - pohjoinen, Keski-, Keski-Länsi, Keski-itä ja Etelä. Kaiken kaikkiaan kaikilla alueilla oli noin 15 tuhatta venäläistä sukunimeä, joista suurin osa löytyi vain yhdeltä alueelta ja puuttui muilta. Kun alueelliset luettelot asetettiin päällekkäin, tutkijat tunnistivat vain 257 niin kutsuttua "kokovenäläistä sukunimeä". Mielenkiintoista on, että tutkimuksen viimeisessä vaiheessa he päättivät lisätä Krasnodarin alueen asukkaiden sukunimet eteläisen alueen luetteloon odottaen, että Katariina II:n täältä häätämien Zaporozhye-kasakkojen jälkeläisten ukrainalaiset sukunimet hallitsevat. , vähentäisi merkittävästi koko Venäjän listaa. Mutta tämä lisärajoitus pienensi kokovenäläisten sukunimien luetteloa vain 7 yksiköllä - 250:een. Tästä seurasi ilmeinen ja ei kaikille miellyttävä johtopäätös, että Kubanissa asui pääasiassa venäläisiä. Ja minne ukrainalaiset menivät ja olivatko ukrainalaisia ollenkaan - iso kysymys.
Kolmen vuoden ajan "Russian Gene Pool" -projektin osallistujat kulkivat ruiskulla ja koeputkella lähes koko Venäjän federaation Euroopan alueen ja tekivät erittäin edustavan näytteen venäläisestä verestä.
Halvat epäsuorat menetelmät venäläisen kansan genetiikan tutkimiseksi (sukunimillä ja dermatoglyfisillä) olivat kuitenkin vain apuväline ensimmäiselle nimellisen kansallisuuden geenipoolin tutkimukselle Venäjällä. Hänen tärkeimmät molekyyligeneettiset tulokset ovat saatavilla monografiassa "Russian Gene Pool" (kustantaja "Luch"). Valitettavasti valtion rahoituksen puutteen vuoksi tutkijat joutuivat suorittamaan osan tutkimuksesta yhdessä ulkomaisten kollegoiden kanssa, jotka asettivat monille tuloksille moratorion, kunnes yhteisiä julkaisuja tieteellisessä lehdistössä julkaistiin. Mikään ei estä meitä kuvailemasta näitä tietoja sanoin. Y-kromosomissa venäläisten ja suomalaisten välinen geneettinen etäisyys on siis 30 tavanomaista yksikköä. Ja geneettinen etäisyys venäläisen ja Venäjän federaation alueella asuvien ns. suomalais-ugrilaisten kansojen (mari, vepsalaiset jne.) välillä on 2-3 yksikköä. Yksinkertaisesti sanottuna ne ovat geneettisesti lähes identtisiä. Mitokondrioiden DNA-analyysin tulokset osoittavat, että tataareista peräisin olevat venäläiset ovat samalla 30 tavanomaisen yksikön geneettisellä etäisyydellä, mikä erottaa meidät suomalaisista, mutta Lvovin ukrainalaisten ja tataarien välillä geneettinen etäisyys on vain 10 yksikköä.... Ja samaan aikaan Ukrainan vasemman rannan ukrainalaiset ovat geneettisesti yhtä lähellä venäläisiä kuin komi-zyryalaiset, mordvalaiset ja marit.
Onko kielillä geenejä? - Miksi geenipoolit tarvitsevat nimiä? - Mitä etäisyyskartat sanovat? - Ei yksi kortti, vaan koko fani!§1. Etäisyydet kolmesta kieliperheestä: INDO-EUROPALAISTA: erot kasvavat itään - Mutta suurin osa populaatioista on geneettisesti läheisiä; - URALISTA: etäisyydet kasvavat idästä länteen - Mutta monet väestöt ovat lähellä - Suomalais-ugrilainen substraatti slaavien ja turkkilaisten keskuudessa; - ALTAISTA: lähellä vain itseään - Ei vaikutusta naapureihin Euroopassa
§2. Etäisyydet venäläisistä, valkovenäläisistä, ukrainalaisista: Klassiset merkit - Pohjoisvenäläiset ovat kauempana keskimääräisistä venäläisistä kuin ukrainalaiset, mordvalaiset ja tšuvashit - Autosomaaliset DNA-merkit - Edellinen kuva - Melkein kaikki ovat lähellä venäläisiä - Kaukasusta ja Uralia lukuun ottamatta - Y-kromosomi - Sama kuva suuremmalla kontrastilla - Etäisyydet valkovenäläisiltä - Samanlainen vain slaaveilla - Sama kuva ukrainalaisilla - Itä-Euroopan väestö on siis lähellä venäläisiä, eikä slaaveja yleensä!
ONKO KIELILLÄ GEENejä?
Haluaisimme vastata heti, että kirjoittajat, kuten lukija, tietävät, että kielillä ei ole geenejä. Tämä on ymmärrettävää jopa arkipäivän tasolla - kuinka monet venäläiset, jotka ovat hajallaan ensimmäisen, toisen ja muun muuttoliikkeen aalloilla, puhuvat erilaisia kieliä! Ja heillä kaikilla on samat geenit, periytyneet esivanhemmiltaan.
Miksi sitten puhumme slaavilaisen tai germaanisen kieliperheen geeneistä? Onko se tieteellinen? Melko. Loppujen lopuksi harjoitamme populaatiogenetiikkaa ja puhumme vain slaavilaisten tai germaanisten kielten kieliä puhuvien ihmisten väestöstä. Eikä "kielisten nimien" takana ole mitään muuta.
Olemme jo todenneet useammin kuin kerran, että populaatiot ovat monikerroksisia ja voivat olla hyvin erilaisia - alkeisväestöstä (useita naapurikyliä) koko ihmiskunnan väestöön. Kaikki nämä ovat populaatioita, ja ne ovat sisäkkäin toistensa sisällä kuin pesimänuket: monet alemman tason populaatiot sopivat seuraavaksi korkeamman tason populaatioon ja niin edelleen. Määrittelemme karkeasti yhden näistä matryoshka-välipopulaatioista etnisen alkuperän mukaan. Vain tästä syystä voimme puhua venäläisestä geenipoolista - eli populaatiosta, jota leimaa ihmisten kuuluminen venäläiseen kansaan. Lisäksi tämän kuulumisen määräävät ihmiset itse, eikä suinkaan genetiikka! Ja vasta kun ihmiset tunnistivat itsensä venäläisiksi tai norjalaisiksi (tai sanoivat, että heidän isovanhempansa ajattelivat sitä), geneetikot alkavat katsoa puolueettomasti: ovatko venäläisten ja norjalaisten populaatiot erilaisia ja kuinka paljon erilaisia? Kutsumme sellaisia populaatioita ehdollisesti "venäläisiksi" tai "norjalaisiksi", samalla kun ymmärrämme täysin, että geenipoolit ja populaatiot ovat biologisia yksiköitä, joille annamme "humanitaarisia" nimiä.
Mutta korostakaamme, koska meillä on nimet "venäläinen" tai "norjalainen" taateleiden geenipankeille, ei tarkoita, että "venäläiset geenit" tai "norjalaiset geenit" olisivat yhtäkkiä ilmestyneet näyttämölle! Ei ole olemassa "venäläisiä" tai "ukrainalaisia" geenejä, kuten ei ole slaavilaisia tai roomalaisia geenejä. Ei, jos vain siksi, että geenit ovat paljon vanhempia kuin ihmiset ja ovat hajallaan melkein kaikkialla maailmassa. Käsittelemme kuitenkin näitä kysymyksiä kirjan lopussa (luku 10). Ja nyt meidän on vain tärkeää vastata kysymykseen - jos venäläisiä tai slaavilaisia geenejä ei ole, miksi kutsumme geenipooleja sellaisiksi nimiksi?
Miksi geenipoolit tarvitsevat nimiä?
Vain siksi, että populaatioille (ja niiden geenipoolille) on annettava selkeät nimet. Voit tietysti jättää geenipoolin nimeämättä ja toistaa koko ajan "Itä-Euroopan tasangon ja pohjoisempien alueiden päämaaseutuväestöä, joka korreloi karkeasti Venäjän valtion rajojen kanssa ennen Ivan Julmaa." ." Mutta tällaisestakin lauseesta jää epäselväksi ketä analysoimme (esimerkiksi otetaanko mukaan karjalaiset, ishoralaiset, tataarit vai mordovialaiset). Ja jos sanomme (kuten kuvailimme yksityiskohtaisesti kirjan alussa), että venäläisellä geenipoolilla tarkoitamme alkuperäiskansoja maaseutuvenäläisiä heidän "alkuperäisellä" (historiallisella) alueella, ja käytämme sitten termiä "venäläinen geenipooli" kauttaaltaan kirja, niin lukijan on helpompi ymmärtää, mistä kirjoittajat puhuvat. Siksi annamme geenipooleille tavanomaisia nimiä - ymmärtämisen helpottamiseksi.
Korkeamman tason matryoshka-nukkejen nimeämiseksi on kuitenkin käytettävä jonkinlaista väestöluokitusta. Luvussa 2 esimerkiksi testasimme, kuinka tehokkaita rotu- ja kielelliset luokitukset ovat geneettisesti. Ja Siperian kansojen keskuudessa tarkastettiin kansojen luokittelun geneettistä tehokkuutta koristetyypin ja shamaanitamburiinien tyypin mukaan. Ja kävi ilmi, että koriste tunnistaa populaatiot huonosti, mutta shamaanitamburiinit eivät ole yhtä tehokkaita populaatioiden tunnistamisessa kuin kielet. Siitä huolimatta kielellinen luokitus on kehitetty yksityiskohtaisimmin. Tästä syystä populaatioiden nimet annetaan usein kielten nimillä. Joten se on nyt hyväksytty biologisissa tieteissä. Ja kun puhumme esimerkiksi geenipoolin "suomalais-ugrilaisesta" kerroksesta, niin antropologit kuin arkeologitkin ymmärtävät meitä. He ymmärtävät, että puhumme tietystä ihmisjoukosta, joka on melko laaja ajassa ja tilassa. Eikä sillä ole väliä, että nyt tšuvashit ovat vaihtaneet aiemman kielensä turkkiksi, ja ei haittaa, ettemme tiedä mitä kieltä muinaiset väestöt puhuivat, jos he eivät jättäneet kirjallisia todisteita. Valtava joukko tietoa eri tieteistä (mukaan lukien esimerkiksi toponyymi - jokien tai järvien nimet) todistaa, että täällä oli ihmisyhteisö, jolle nyt annamme väliaikaisen nimen "suomalais-ugrilaiseksi" maailmaksi.
Siksi sekä tässä että seuraavassa osiossa, jossa verrataan geneettisiä etäisyyksiä populaatioista, joilla on "kieliset" nimet, emme petä tiedettä, vaan noudatamme sen tiukkoja sääntöjä. Otamme kansojen kielellisen luokituksen; sitten sen mukaisesti annamme jokaiselle populaatioryhmälle tavanomaisen "kielisen" nimen; ja lopuksi laskemme keskimääräiset geenifrekvenssit niille tämän ryhmän populaatioille, jotka elävät analysoidulla alueella. Ja sitten katsomme, kuinka kukin Itä-Euroopan väestö eroaa näistä "indoeurooppalaisten" tai "altailaisten" väestön keskimääräisistä taajuuksista. Samanaikaisesti kirjoittajat, kuten lukija, tietävät, että Altailaisia kieliä Euroopassa puhuvat ihmiset, joilla on täysin erilainen fyysinen ulkonäkö - gagauzista kalmykiin. Mutta meillä ei ole tällä perusteella oikeutta sulkea ketään pois niistä ryhmistä, jotka kielitiede on tunnistanut - luettelemme rehellisesti, mitkä kansat kuuluvat väestöön tietyllä "kielitieteellisellä" nimellä.
MITÄ ETÄISYÖKORTIT KERROVAT?
Geneettiset etäisyyskartat ovat melkein tärkeämpiä kuin pääkomponenttikartat. Nämä kaksi genogeografista perustyökalua, yhdessä käytettynä, tarjoavat täydentävän kuvauksen geenipoolista. Pääkomponenttikarttojen avulla voimme esittää hypoteeseja tekijöistä, jotka muodostivat havaitut kuviot, ja geneettisten etäisyyskarttojen avulla voimme testata näitä hypoteeseja.
Jokainen tässä osiossa annettu geneettisten etäisyyksien kartta on kaikkien tutkittujen lokusten keskiarvo (taulukko 8.1.1.). Se osoittaa selvästi, kuinka geneettisesti lähellä kukin alueen populaatio on tutkijan määrittelemää populaatioryhmää. Tällaista populaatioiden ryhmää kutsutaan "viiteeksi".
Geenipoolilta voidaan esittää kysymyksiä: mitkä populaatiot ovat geneettisesti lähellä meitä kiinnostavaa väestöryhmää? Mitkä ovat suhteellisen etäisiä? Ja mitkä niistä poikkeavat perustavanlaatuisesti vertailuryhmästä, lisäksi koko geenitaajuuksien joukossa? Ja geneettisten etäisyyksien kartta antaa vastauksen: kuinka paljon jokainen kartan piste on geneettisesti lähellä tai kaukana vertailuryhmästä. Näemme sen omin silmin.
Etäisyyskartat poikkeavat tavanomaisesta geneettisten etäisyyksien käytöstä vain yhdellä, mutta tärkeimmällä ominaisuudella: kartoittaessa analyysi sisältää väestön alueen eli maantieteellisen, spatiaalisen näkökulman.
Geneettiset etäisyyskartat paljastavat usein geneettisten ja maantieteellisten etäisyyksien välisen suhteen. Kartta osoittaa, kuinka etäisyyden (tutkijan antama) vertailupopulaatiosta viereisten ja syrjäisten alueiden populaatio muuttuu geneettisesti yhä enemmän erilaiseksi kuin vertailupopulaatio. Tämä geneettisen etäisyyden kasvu ei kuitenkaan riipu vain maantieteellisestä etäisyydestä. Muuten mikä tahansa geneettisten etäisyyksien kartta koostuisi samankeskisistä ympyröistä, jotka ovat samanlaisia kuin ne, jotka leviävät veteen heitetyistä kivestä.
Todellisuudessa etäisyydet yhteen suuntaan voivat kuitenkin kasvaa nopeasti, mikä osoittaa geenivirran esteitä; muihin suuntiin etäisyydet saattavat tuskin kasvaa, mikä osoittaa näiden vierekkäisten ryhmien geneettisen affiniteetin. Monissa tapauksissa isoliinien sujuva kulku voi häiriintyä ja geneettisesti läheisten ryhmien joukosta tunnistetaan geneettisesti etäinen populaatio, mikä voi viitata esimerkiksi sen muuttoon tälle alueelle. Siten geneettisten etäisyyksien piirtäminen kartalle voi antaa arvokasta tietoa tutkitun ryhmän suhteesta alueen muihin populaatioihin, geneettisten virtojen esiintymisestä, geneettisistä esteistä ja niihin liittyvistä ryhmistä. Lisäksi saamme tietoa itse vertailuryhmästä (esimerkiksi venäläisistä tai valkovenäläisistä): geneettisestä monimuotoisuudesta sen rajoissa, poikkeamista keskiarvoista sen omalla alueella.
EI YKSI KARTTA. KOKO FANI!
Geneettisten etäisyyksien kartoitus selventää monia geenipoolin piirteitä - varsinkin jos otamme huomioon ei yhden etäisyyskartan (yhdestä ihmisestä), vaan sarjan karttoja - eri kansoista, suurista väestöryhmistä. Jokainen uusi kartta kertoo uuden kansan tai kansanryhmän geneettisestä asemasta alueen yleisessä geenipoolissa. Koko karttafanien vertailu osoittaa, kuinka suuri kunkin näistä ryhmistä on panos Itä-Euroopan geenipooliin ja missä niiden sekoittumisvyöhykkeet sijaitsevat.
Emme käsittele tässä kaikkien Itä-Euroopan ihmisten geneettisten etäisyyksien karttoja - joten menisimme liian pitkälle venäläistä geenipoolia käsittelevän kirjan soveltamisalan ulkopuolelle. Informatiivisempia karttoja etäisyyksistä ihmisryhmistä, jotka liittyvät toisiinsa. Ne eivät paljasta yksittäisten kansojen etnogeneesin malleja, vaan Itä-Euroopan väestön muodostumisen yleisiä tapahtumia. Kuten luvussa 2 totesimme, genogeografia perustuu "skaalauksen" periaatteeseen: tutkittavien ryhmien mittakaavan kasvaessa paljastuu jälkiä vanhemmista ja laajemmista tapahtumista.
Siksi huomiota kiinnitetään karttoihin etäisyyksistä ihmisryhmistä. Kohdassa 1 kartat on rakennettu Itä-Euroopassa asuvien indoeurooppalaisten, uralilaisten ja altailaisten kieliperheiden kansojen geenien keskimääräisistä frekvenssistä. Sitten (§2) tarkastellaan karttoja etäisyyksistä venäläisistä, jotka osoittavat sen aseman Itä-Euroopan yleisessä geenipoolissa. Ja lopuksi, katsotaanpa kahden muun Itä-Euroopan kansan - valkovenäläisten ja ukrainalaisten - karttoja, jotka ovat historiallisesti lähellä venäläisiä populaatioita ja joilla voi olla samanlainen geenipooli.
Kaikki kortit luetaan samalla tavalla. Mitä enemmän tietty kartan piste on geneettisesti poistettu vertailupopulaatiosta, sitä suurempi on etäisyys, sitä voimakkaampi tämän pisteen väri on. Siksi kevyimmät alueet ovat lyhimpien etäisyyksien alueita. Nämä ovat populaatiot, jotka muistuttavat eniten vertailuarvoa. Tummin ovat alueet, jotka ovat suurimman etäisyyden päässä. Nämä ovat populaatioita, jotka eivät ole geneettisesti samanlaisia kuin vertailupopulaatio. Tietysti. heti kun otamme erilaisen vertailupopulaation, samat kartan pisteet ilmoittavat, että niillä on jo eri etäisyydet uuteen referenssiin. Lukemisen helpottamiseksi kaikki etäisyyskartat on rakennettu samaan mittakaavaan, joten voit turvallisesti verrata saman kartan eri osia, mutta myös eri karttoja keskenään värien voimakkuuden suhteen.
§1. Etäisyydet kolmesta kieliperheestä
Harkitse karttoja kaikkien Itä-Euroopan populaatioiden geneettisistä etäisyyksistä siellä asuvista indoeurooppalaisten, uralilaisten ja altailaisten kieliperheiden kansoista. Esitämme lyhyyden vuoksi karttoja yhdestä "silminnäkijästä" - autosomaalisista DNA-markkereista, koska klassisten markkerien geneettisten etäisyyksien kartat, kuten seuraavassa osiossa näemme, ovat melko samanlaisia.
INDO-EUROPAN KIELIPERHEEN KANSALTA (DNA-MERKKIT)
Kartta geneettisistä etäisyyksistä indoeurooppalaisesta kieliperheestä on esitetty kuvassa. 8.3.1.
Kartta rakennettiin näin. Ensin laskettiin DNA-merkkien keskimääräiset taajuudet Itä-Euroopan indoeurooppalaisen perheen edustajille: venäläisten, ukrainalaisten, valkovenäläisten ja moldovalaisten populaatioille. Sitten niiden perusteella saatiin keskimääräiset "indoeurooppalaiset" geenitaajuudet. Lisäksi lasketaan geneettiset etäisyydet näistä keskimääräisistä "indoeurooppalaisista" taajuuksista kartan kunkin pisteen taajuuksiin ja saadut etäisyysarvot sijoitetaan kartan samoihin solmuihin.
Siksi, jos esimerkiksi suurimmassa osassa Valko-Venäjää, Kiovan ja Lvovin alueilla geneettisten etäisyyksien arvot putoavat välillä 0,01 - 0,02 (kuva 8.3.1.), Tämä tarkoittaa, että nämä ovat ( keskimäärin kaikkien geenien osalta) näiden populaatioiden väliset erot indoeurooppalaisen perheen kansojen keskimääräisistä esiintymistiheyksistä. Päinvastoin, erot kalmykkien, komien ja baškiirien välillä ovat paljon suuremmat - geneettisten etäisyyksien arvot heidän asutusalueillaan ovat yli 0,05 ja 0,06. Loput geneettisten etäisyyksien kartat luetaan samalla tavalla.
Kartta osoittaa, että Keski-Venäjän venäläisten, ukrainalaisten, valkovenäläisten ja moldovalaisten (eli itse indoeurooppalaisten väestön) populaatiot ovat, kuten arvata saattaa, lähellä Itä-Euroopan indoeurooppalaisten kansojen keskimääräisiä esiintymistiheyksiä. . Kaikki ei kuitenkaan ole ennustettavissa - pohjoisen venäläiset väestöt (vaikka he ovat myös indoeurooppalaisia) eroavat huomattavasti "keskimääräisistä indoeurooppalaisista" - samassa määrin kuin Keski-Volgan ei-indoeurooppalaiset kansat (mari, mordvalaiset, tšuvashit) ja Länsi-Kaukasus. Lopuksi erinomaisin on Uralin väestö (erityisesti komit), samoin kuin arokansat (baškiirit, kalmykit).
Kiinnitämme huomiota Venäjän kansan väestöön. Ne edustavat indoeurooppalaista kieliperhettä Itä-Euroopassa, niiden taajuuksia käytettiin keskimääräisten "indoeurooppalaisten" taajuuksien laskemiseen. Ja siitä huolimatta näemme Venäjän väestön silmiinpistäviä eroja oman vertailuväestön läheisyyden asteessa. Tämä taas osoittaa, että venäläisten geenipoolin heterogeenisyyden taso on niin suuri, että se näkyy selvästi jopa Itä-Euroopan mittakaavassa.
Yleisesti ottaen paljastuu selvä maantieteellinen kuvio: itään siirryttäessä etäisyysarvot kasvavat vähitellen, populaatiot poikkeavat yhä enemmän indoeurooppalaisten kansojen ja itäisten esikaupunkien kansojen keskimääräisistä ominaisuuksista. Eurooppa osoittautuu geneettisesti kaikkein erilaisimmaksi kuin he. Yleisesti ottaen suurin osa Itä-Euroopan kansoista (mukaan lukien Ural- ja Kaukasian väestöt mukaan lukien) ovat kuitenkin lähellä indoeurooppalaisia: geneettisten etäisyyksien keskiarvo kartalla on pieni d = 0,028.
URALIN KIELIPERHEEN KANSSA (DNA-MERKKIJÄ)
Seuraava geneettisten etäisyyksien kartta on rakennettu uralilaisen kieliperheen geenien keskimääräisistä taajuuksista ja näyttää toisenlaisen kuvan (kuva 8.3.2.).
Uralin suvusta vain itäsuomenkielisiä kansoja (komit, udmurtit, marit, mordvalaiset) on tutkittu DNA-merkkien avulla. Vähimmäisetäisyydet löytyvät näiden kansojen asutusalueelta, pääasiassa Uralilta. Päinvastoin, Venäjän tasangon ja Ciscaukasian länsiosan väestö on geneettisesti kaukana Uralin keskimääräisistä taajuuksista. Itä-Euroopan keskialueet, maantieteellisesti Uralin vieressä, ovat lähempänä uralilaisia ja geneettisesti.
Niin, etäisyyksien pienimmät arvot sijaitsevat Uralissa ja kasvavat vähitellen edelleen länteen. Todennäköisesti väliarvojen miehittämät alueet heijastavat slaavien assimiloimien muinaisten suomalais-ugrilaisten heimojen aluetta [Alekseeva, 1965]. On uteliasta, että Uralin turkinkielisten kansojen alueet ovat lähellä uralilaisten suvun piirteitä, mikä selittyy uralilaisen substraatin merkittävällä osuudella tšuvashien, tataarien ja joidenkin ryhmien geenipoolissa. Baškiirit [Roginsky, Levin, 1978].
Keskimääräinen etäisyys kartalla on pieni (d = 0,039), vaikka se onkin suurempi kuin "indoeurooppalaisen". Tämä vahvistaa uralinkielisen geenipoolin merkittävän edustuksen yleisessä Itä-Euroopan geenipoolissa, joka koostuu suurelta osin uralilaisesta substraatista.
ALTAIN KIELIPERHEEN KANSASTA (DNA-MERKKEJÄ)
Seuraava kartta (Kuva 8.3.3.) osoittaa erot kunkin Itä-Euroopan väestön ja Altain kieliperheen kansojen välillä. Tätä Itä-Euroopan perhettä edustavat pääasiassa turkkia puhuvat kansat - vain kalmykit puhuvat kieltä, joka kuuluu tämän perheen mongoliryhmään.
Kahdelle aikaisemmille geneettisten etäisyyksien kartalle (indoeurooppalaisesta ja uralilaisperheestä) oli ominaista etäisyyksien pienet keskiarvot. Kartoissa (kuva 8.3.1., 8.3.2.) tämä havaittiin vaaleiden sävyjen vallitsevana. Päinvastoin, Altai-suvun etäisyyskartalla (kuva 8.3.3.) vallitsee tumma väri, joka vastaa useimpien Itä-Euroopan populaatioiden merkittävää geneettistä etäisyyttä Altain kieliperheen geenipoolista. Vain Altain kieliperheen kansojen itsensä alueet ovat luonnollisesti lähellä keskiarvojaan. Ja heti asutusalueensa ulkopuolella muut Itä-Euroopan populaatiot osoittautuvat jyrkästi erilaisiksi Altain puhuvien kansojen geneettisistä ominaisuuksista.
Tämä näkyy geneettisten etäisyyksien suurempana arvona kuin aikaisemmissa kartoissa. Keskimäärin kartalla ne olivat d = 0,064, mikä on lähes kolme kertaa suurempi kuin indoeurooppalaisten kansojen vastaava arvo.
Siten Altai-perheen kansojen vaikutusta Itä-Euroopan geenipooliin rajoittaa vain heidän asutusvyöhyke, ja tarkasteltavien tietojen mukaan sitä ei käytännössä jäljitetä edes viereisillä alueilla. Tämä tosiasia voidaan selittää sillä, että monet heimot ilmaantuivat suhteellisen myöhään Itä-Eurooppaan, jotka puhuvat Altai-suvun kieliä [Maailman kansakunnat ja uskonnot, 1999], kun taas sekä indoeurooppalaiset että uralilaiset perheet ovat kieliä. Itä-Euroopan vanhempi väestö [Cheboksarov, Cheboksarova, 1971; Bunak, 1980].
§2. Etäisyydet venäläisistä, valkovenäläisistä, ukrainalaisista
Olemme siis oppineet Itä-Euroopan geenipoolin pää"koostumuksen" - mitkä ovat sen tärkeimmät alageenipoolit, missä "osuuksissa" ne "sekoittuvat" ja miten nämä osuudet eroavat Itä-Euroopan eri osissa. Nyt voimme palata kirjamme pääaiheeseen ja pohtia, mikä on kaikkien Itä-Euroopan väestön asema venäläisiin nähden? Koska tämä on johtava aihe, annamme geneettiset etäisyydet Venäjän populaatioista kolmen tyyppisille markkereille - klassisille markkereille, autosomaalisille DNA-markkereille ja Y-kromosomin markkereille. Ja jotta "puhtaasti venäläisiä" ja "slaavilaisia" ei sekoitettaisi, harkitsemme myös karttoja etäisyyksistä historiallisesti läheisistä itäslaavilaisista kansoista - valkovenäläisistä ja ukrainalaisista.
VENÄJÄN VÄESTÖSTÄ (KLASSISET MERKINNÄT)
Geneettisten etäisyyksien kartta Venäjän keskimääräisistä taajuuksista klassisilla markkereilla osoittaa kunkin Itä-Euroopan populaation samankaltaisuuden asteen Venäjän geenipoolin kanssa. Keski-Venäjän geenitaajuuksia lähimpänä oleva valoalue sijaitsee Itä-Euroopan keskikaistalla - Valko-Venäjältä Volgan keskialueelle (kuva 8.3.4.). Tummat sävyt ovat alueita, jotka ovat geneettisesti kaukana venäläisistä. Niitä on suhteellisen vähän - etäisyyden mukaan keskivenäläisistä - nämä ovat Krim ja Mustanmeren alue, Ala-Volga, Baltian maat, Venäjän pohjoisosa, Fennoskandia ja geneettisesti kaukana olevat Uralit.
Valkovenäjän ja ukrainalaisten elinympäristöt osoittavat yhtäläisyyksiä Venäjän geenipoolin kanssa. Venäjän pohjoisen ja koillis-Euroopan jyrkät geneettiset erot yleensä, mukaan lukien Vjatka, muinainen Novgorodin siirtokunta, ovat yllättäviä.
Tietenkin näillä alueilla elävä alkuperäisväestö kantaa nyt suurimmassa määrin assimiloituneen väestön piirteitä. On kuitenkin uskomatonta, että suomalais-ugrilaisen väestön osuus täällä oli suurempi kuin mordvalaisten ja tšuvashien populaatioissa, jotka kartalla kuuluivat kokonaan "Keski-Venäjän geneettiseen alueeseen". Tällaisten erojen mahdollista lähdettä on kolme. Ensinnäkin suomalais-ugrilainen substraatti voi itse vetää läntisen suomenkielisten kansojen puoleen, ei itäisiin.
Toiseksi, kuten arkeologiset tiedot osoittavat [Sedov, 1999], Novgorodin kolonisaatiolla oli erilainen alkuperä slaavilaisten heimojen kannalta. Tämä tarkoittaa, että ei vain substraatti, vaan myös slaavilainen superstratum voisi olla ainutlaatuinen Venäjän pohjoisosassa. Kolmanneksi pienissä pohjoisissa populaatioissa geeniryömintätekijä on voimakkaampi, mikä voisi myös "kantaa" ne pois Venäjän päämantereelta. Todennäköisesti kaikki kolme tekijää toimivat rinnakkain, mutta tulevan tutkimuksen tehtävänä on selvittää niiden todellinen suhde. Tässä homogeeniset markkerit voivat olla suureksi avuksi, mikä auttaa erottamaan muuttovirrat avaruudessa ja ajassa.
"Keskivenäläisten" taajuuksien läheisyys ilmenee Venäjän alueen eri osissa, myös niissä, joissa Itä-Euroopan geenipoolin pääkomponenttien vastakkaiset ääripäät sijaitsevat (luku 8.2.). Samankaltainen kuva voidaan selittää hypoteesin perusteella, että "Keski-Venäjän" taajuudet ovat itse asiassa "keskeurooppalaisia" ja venäläinen geenipooli muodostuu monenlaisten itäeurooppalaisten komponenttien sekoituksista (suomalais-ugrilaiset, slaavilainen, balttilainen jne.). Tämä hypoteesi vahvistetaan myös ukrainalaisten, valkovenäläisten ja venäläisten geneettisten etäisyyksien kartoissa informatiivisimman DNA-markkerin - Y-kromosomin haploryhmien - mukaan.
VENÄJÄLLISESTÄ VÄESTÖSTÄ (AUTOSOMISET DNA-MERKKIT)
Kuten klassisten markkerien tietojen mukaan (kuva 8.3.4), Keski-Venäjän populaatio on jälleen lähellä Keski-Venäjän geenitaajuuksia (kuva 8.3.5.). Valkovenäjät, jotka eivät käytännössä eroa keskimääräisistä venäläisistä ominaisuuksista klassisten markkerien taajuuksien suhteen ja osoittavat DNA-tietojen mukaan pieniä eroja. Uralin, Kaukasuksen, Volgan alueen ja vähemmässä määrin Venäjän pohjoisosan populaatiot eroavat suuresti Venäjän keskimääräisistä taajuuksista. Siten kaikissa pääkohdista DNA:n ja klassisten markkerien käyttö johtaa samanlaisiin tuloksiin. Erot näiden kahden kartan välillä johtuvat mielestämme pääasiassa erityyppisten markkerien tuntemusasteesta, ja on odotettavissa, että DNA-polymorfismitietojen kertyessä niiden vaihtelumalli lähestyy yhä enemmän käyttämällä saatuja tuloksia. klassiset merkit.
Itä-Euroopan populaatioiden keskimääräinen geneettinen etäisyys venäläisistä taajuuksista on pieni (d = 0,28), mikä voi johtua venäläisen geenipoolin pitkäaikaisesta vuorovaikutuksesta ympäristön kanssa. Muista, että etäisyydelle indoeurooppalaisista kansoista yleensä on ominaista sama keskiarvo (d = 0,28). Näitä karttoja verrattaessa (kuva 8.3.1. ja 8.3.5) niiden merkittävä samankaltaisuus käy ilmi. Tämä on ymmärrettävää, sillä venäläiset ovat myös indoeurooppalaisia ja taajuudet venäläisväestössä otettiin mukaan laskelmaan indoeurooppalaisten kansojen osalta. On kummallista, että indoeurooppalaisten kansojen keskitaajuuksien etäisyyksien karttaan merkityt erot Venäjän väestön välillä Volgan ja Vyatkan välillä ovat säilyneet myös etäisyyskartalla Venäjän keskitaajuuksista.
Joten Venäjän geenipooli osoittautuu olevan kiinteästi yhteydessä monien Itä-Euroopan kansojen geenipooleihin - geenitaajuuksien suhteen valkovenäläiset, ukrainalaiset, mordvalaiset ja monet muut Itä-Euroopan populaatiot ovat erittäin lähellä venäläisiä. Vasta kun lähestymme Kaukasusta ja Uralia, populaation geenipooli eroaa selvästi Venäjän geenipoolin keskimääräisistä ominaisuuksista. Tämä tulos ei ole odottamaton, koska Venäjän asettuminen laajoille alueille ja intensiivinen geenien vaihto "alkuperäisen" alueen ulkopuolella ympäröivien kansojen kanssa on ilmeistä. Mielenkiintoiselta vaikuttaa pikemminkin se, että genogeografisissa kartoissa esiintyi kaksi vuoristoestettä (Kaukasus ja Ural), mikä rajoitti jossain määrin tätä geenipoolin alueellista laajenemista.
VENÄJÄLLISESTÄ VÄESTÖSTÄ (Y-KROMOSOMIT)
Tässä kortissa on kaksi ominaisuutta. Ensinnäkin, siinä näemme koko Euroopan, eikä vain sen itäpuoliskoa (kartta on rakennettu niiden kahdeksan yksittäisten haploryhmien kartan pohjalta, joita tarkasteltiin kohdassa 6.3). Toiseksi, Y-kromosomimarkkerien erilaistumiskyky on paljon korkeampi, joten erot venäläisten populaatioiden ja naapureiden välillä ovat selvempiä. "Laajemmasta" intervalliasteikosta huolimatta maksimietäisyyksien väli hallitsee karttaa - Y-kromosomimarkkerien mukaan lähes koko Eurooppa eroaa merkittävästi Venäjän geenipoolista (kuva 8.3.6). Vain venäläiset ja valkovenäläiset itse ovat lähimpänä Venäjän keskimääräistä taajuutta, ukrainalaiset, länsislaavit (puolalaiset, tšekit, slovakit) ja Volgan alueen kansat osoittavat keskimääräistä läheisyyttä. Kuten aikaisemmissa kartoissa, pohjoisen Venäjän populaatiot osoittavat selkeää erottuvuutta, joka eroaa jyrkästi keskimääräisestä venäläisestä geenipoolista.
Näemme, että Y-kromosomin markkerit vahvistavat aiemmin tunnistetut "keskivenäläisen" geenipoolin samankaltaisuusmallit muiden itäslaavilaisten kansojen ja Volgan alueen kansojen kanssa sekä Venäjän pohjoisen erot. Y-kromosomin korkea informaatiosisältö tekee näistä kuvioista kuperampia kuin muun tyyppisissä markkereissa, ja koko Euroopan mittakaavassa huomioiminen lisää venäläisen kansojen ja puolalaisten geenipoolin kaltaisten kansojen listaa.
VALKOVENÄJÄLTÄ (KLASSISET MERKKIT)
Aiemmissa kartoissa (kuvat 8.3.4., 8.3.5., 8.3.6.) Näimme, että monet Itä-Euroopan populaatiot ovat samanlaisia kuin Venäjän geenipooli.
On tärkeää ymmärtää: ovatko kaikki nämä populaatiot lähellä Venäjän geenipoolia vai monia itäslaavilaisia populaatioita? Toisin sanoen: onko tämän samankaltaisuuden salaisuus venäläisen kansan etnisessä historiassa tai itäslaavien laajenemisessa kokonaisuudessaan ja mahdollisesti slaavilaisen ja suomalais-ugrilaisen geenin "alkuperäisessä", ennen laajentumista altaat?
Vastataksemme tähän kysymykseen analysoimme Itä-Euroopan geenipoolin läheisyyttä valkovenäläisille - toiselle itäslaavilaiselle etnoselle, joka on maantieteellisesti, etnogeneesistään ja antropologisesti hyvin lähellä venäläisiä.
Kuvassa 8.3.7. kartta Itä-Euroopan populaatioiden geneettisistä etäisyyksistä Valko-Venäjän keskimääräisistä geenitaajuuksista suurelle joukolle klassisia geenimarkkereita - esitetään 21 lokuksen 57 alleelia. Näemme selkeän kuvan, joka poikkeaa olennaisesti Venäjän geenipoolin vaihtelevuuden luonteesta. Lähes kaikki alueet, joiden väestö on lähimpänä Valko-Venäjän geenipoolia, sijaitsevat Valko-Venäjän alueella. Valko-Venäjän alueen ulkopuolella geneettiset etäisyydet kasvavat nopeasti merkittäviin arvoihin, mikä viittaa selkeisiin geneettisiin eroihin valkovenäläisten geenipoolin ja Itä-Euroopan geenipoolin välillä kokonaisuudessaan.
Kartta vangitsee valkovenäläisten geenipoolin geneettisen omaperäisyyden, mikä kertoo geneettisen etäisyyden menetelmän korkeasta herkkyydestä. Huomaa, että selkeät erot Valko-Venäjän geenipoolin ja naapurialueiden geenipoolin välillä ovat tärkeä odottamaton tulos, koska antropologiset tiedot eivät yleensä paljasta selkeitä eroja valkovenäläisten ja naapuriryhmien välillä [Alekseeva, 1973; Deryabin, 1999]. Tietenkin tämä valkovenäläisten geneettinen erityispiirre on hyvin suhteellinen: se ilmenee vain Valko-Venäjän mittakaavassa, ikään kuin mikroskoopin läpi, kiitos karttojen valtavan resoluution, jotta voidaan nähdä jopa hienovaraisia yksityiskohtia. Muista, että eri mittakaavassa - venäläisten geneettisten etäisyyksien kartoissa - valkovenäläisiä ei käytännössä voida erottaa Keski-Venäjän venäläisistä. Joka tapauksessa valkovenäläiset ovat paljon enemmän heidän kaltaisiaan kuin Venäjän pohjoisen venäläiset itse.
Siten, toisin kuin venäläinen, Valko-Venäjän geenipooli ei ole lähellä Itä-Euroopan geenipoolia kokonaisuutena. Näin ollen venäläisten populaatioiden suuri geneettinen samankaltaisuus useimpien Itä-Euroopan alueiden väestön kanssa ei ole kaikille itäslaavilaisille kansoille yhteinen piirre, vaan sen oma ominaisuus Venäjän geenipoolissa.
VALKOVENÄJÄLTÄ (MARKERS Y KROMOSOMES)
Tämän päätelmän vahvistavat Y-kromosomista saadut tiedot. Etäisyyskartta valkovenäläisistä (kuva 8.3.8.) on rakennettu samaan mittakaavaan kuin venäläisiltä (kuva 8.3.6.). Mutta vyöhyke, joka on geneettisesti samanlainen kuin Valko-Venäjän geenipooli, on huomattavasti pienempi: se sisältää vain slaavilaisia kansoja (sekä itäslaavia, paitsi Länsi-Ukrainaa, että länsislaavialaisia populaatioita), mutta se ei sisällä Volgan ja Uralin alueiden kansoja. Siten geneettinen yhteisyys Itä-Euroopan ei-slaavilaisten väestöjen kanssa on Venäjän geenipoolin "etuoikeus", toisin kuin valkovenäläisten geenipooli, joka eroaa jyrkästi näistä Volgan alueen ja Uralin kansoista.
UKRAINAlaisilta (MARKERS Y KROMOSOMES)
Itäslaavilaisten kansojen tarkastelun täydellisyyden vuoksi esittelemme myös kartan etäisyyksistä ukrainalaisista (kuva 8.3.9.). Se on hyvin samanlainen kuin valkovenäläisiltä juuri tarkasteltu kartta, vain suurimman läheisyyden vyöhyke on siirretty itse ukrainalaisten alueelle, ja tämä vyöhyke sisältää myös eteläisen venäläisen ja valkovenäläisen väestön. Ja Itä-Euroopan ei-slaavilaiset kansat, jotka ovat suhteellisen lähellä venäläisiä väestöjä, ovat yhtä kaukana ukrainalaisten geenipoolista kuin valkovenäläisten geenipoolista. Tämä vahvistaa tulkintamme oikeaksi, että Itä-Euroopan tasangon slaavilaiseen kolonisaatioon, johon liittyi suomalais-ugrilaisen väestön assimilaatio, osallistuivat pääasiassa nykyisen venäläisen väestön esi-isät koko slaavilaismassivista.






Tutkijat ovat luoneet ensimmäistä kertaa yksityiskohtaisen kartan Yhdistyneen kuningaskunnan populaatioiden geneettisestä rakenteesta. Kävi ilmi, että kelttiläisen alueen populaatio ei ole geneettisesti ollenkaan homogeenista, ja vanhin geenipooli on säilynyt Walesissa. Brittien ja mannereurooppalaisten DNA:n vertailu osoitti kunkin alueen esi-isien geneettisen profiilin. Nykyajan väestön geenipoolissa oli mahdollista nähdä heijastus Brittisaarten asutushistorian tärkeimmistä tapahtumista.
Nykyajan populaatioiden geneettisten erojen tutkiminen auttaa katsomaan syvälle historiaan ja näkemään jälkiä väestöryhmien muuttoliikkeestä, jonka ansiosta nykyaikainen geenipooli muodostui. Tämän työn suoritti kansainvälinen tutkijaryhmä, jota johti brittiläiset asiantuntijat Oxfordin yliopistosta ja University College Londonista, heidän artikkelinsa julkaistiin Nature-lehdessä 19. maaliskuuta. Walter Bodmer, Peter Donnelly ja heidän kollegansa analysoivat yksityiskohtaisesti Yhdistyneen kuningaskunnan väestön geneettistä rakennetta. He eivät vain rakentaneet erittäin tarkan geneettisen kartan nykyaikaisesta brittiläisestä väestöstä, vaan löysivät myös jälkiä tärkeimmistä historiallisista muuttoista ja selvensivät Brittisaarten asutuksen luonnetta.
Geenivarasto heijastaa maantiedettä
Tutkijat keräsivät DNA:ta 2 039 ihmiseltä eri puolilta Isoa-Britanniaa. Valitessaan heitä ohjasi seuraava sääntö: kaikkien kolmannen sukupolven ihmisten esivanhempien (sekä isoäitien että molempien isoisien) oli asuttava enintään 80 km päässä toisistaan. Tämä ehto mahdollisti "tilannekuvan" Britannian väestön geneettisestä rakenteesta 1800-luvun lopulla ja 1900-luvun alussa.
DNA:n vertaamiseen tutkijat käyttivät yli 500 000 yhden nukleotidin polymorfismia (SNP:tä), jotka olivat hajallaan genomissa. Niiden analyysi suoritettiin tilastollisilla menetelmillä fineSTRUCTURE ja GLOBETROTTER. Nämä menetelmät luokittelivat DNA-näytteet klustereihin niiden samankaltaisuuden perusteella riippumatta siitä, mistä ne on kerätty Yhdistyneessä kuningaskunnassa. Ja asettamalla näytteen jokaisesta henkilöstä puoliväliin isovanhempiensa syntymäkoteja, he pystyivät näkemään, kuinka geneettinen samankaltaisuus korreloi maantieteellisen sijainnin kanssa.
Geneettiset klusterit, jotka ryhmittelivät 2039 DNA-näytettä Yhdistyneen kuningaskunnan populaatioista.
Sattuma oli silmiinpistävä. Kuten kuvasta näkyy, 17 geeniklusteria jakaantui kartalle otettujen näytteiden maantieteellisen sijainnin mukaisesti. Eri klusterit eivät käytännössä mene päällekkäin. Ja koska nämä klusterit muodostettiin kaikista näytteistä ottamatta huomioon niiden maantieteellistä sijaintia, tämä tarkoittaa, että tämä vastaavuus heijastaa populaation todellista geneettistä monimuotoisuutta, kirjoittajat korostavat.
Skotlannin pohjoispuolella sijaitsevien Orkneysaarten Orkneyn piirikunnan väestö osoittautui geneettisesti monimuotoisimmaksi, se sisältää kolme klusteria. Geneettiset erot ovat selviä kaikkialla Yhdistyneessä kuningaskunnassa: Pohjois-Englanti, Skotlanti ja Pohjois-Irlanti eroavat kollektiivisesti Etelä-Englannista, Pohjois-Wales on erillään Etelä-Walesista ja Cornwall muodostaa erillisen klusterin. Klusterien rajat osuvat usein yhteen historiallisten provinssien nykyaikaisten rajojen kanssa, esimerkiksi Cornwallin ja Devonin rajalla, Englannin ja Walesin rajalla. Suurin klusteri (merkitty punaisilla neliöillä kartalla) kattaa suurimman osan Keski- ja Etelä-Englannista sekä itärannikosta. Se yhdistää lähes puolet tutkimukseen sisältyvistä DNA-näytteistä.
Kirjoittajat rakensivat brittiläisistä populaatioista fylogeneettisen puun, joka näyttää kuinka klustereita muodostui, kuinka pääoksat erotettiin. Kuten puusta näkyy, varhaisimmassa vaiheessa Orkneysaarten väestön haara erottui, seuraavassa vaiheessa Wales. Myöhemmät haarukat puussa johtivat Pohjois- ja Etelä-Walesin erottamiseen, Pohjois-Englannin, Skotlannin ja Pohjois-Irlannin erottamiseen muusta Englannista ja Cornwallin erottamiseen muista klusteista.
Siten ajatusta Ison-Britannian kelttiläisten populaatioiden geneettisestä homogeenisuudesta vanhimpana, johon Saksien vaellus ei vaikuttanut, ei vahvistettu. Päinvastoin, Ison-Britannian kelttiläinen osa (Skotlanti, Pohjois-Irlanti, Wales ja Cornwall) osoittautui geneettisesti paljon monimuotoisemmaksi kuin muu alue.
Vaikka brittiväestön sisällä olevat klusterit erottuivat niin selvästi, tutkijoiden välinen geneettinen etäisyys osoittautui pieneksi (keskiarvo 0,002, maksimi 0,007). Tämä tarkoittaa, että havaitut erot ovat hyvin hienovaraisia ja ne voidaan nähdä vain sillä vahvalla "suurennuksella", jonka tutkijat loivat tehokkaalla tilastollisella menetelmällä. Vertailun vuoksi he analysoivat tietojaan standardimenetelmillä - pääkomponenttianalyysillä (PCA) ja ADMIXTURE:lla. Molemmat menetelmät osoittivat oksien erottamisen Orkneysta ja Walesista, mutta eivät paljastaneet populaation hienompaa geneettistä rakennetta.
Manner jalanjälki brittiläisessä genomissa
Geneettiset erot brittiväestön sisällä voivat johtua tiettyjen väestöryhmien eristäytymisestä toisistaan, ja lisäksi ne heijastavat erilaisten muuttoliikkeiden osuutta ja siirtolaisten sekoittumista alkuperäiskansojen kanssa. Viimeksi mainitun ratkaisemiseksi tutkijat käyttivät samaa menetelmää vertaillakseen DNA-näytteitä Britannian väestöstä yli 6 000 mannereurooppalaisen näytteisiin 10 maasta. Geneettisen samankaltaisuuden mukaan nämä näytteet jaettiin 51 ryhmään.

Eurooppalaiset geneettiset profiilit 17 Yhdistyneen kuningaskunnan klusterille.
Kunkin brittiläisen klusterin osalta tutkijat arvioivat kunkin mannerryhmän panoksen ja määrittelivät esi-isiensä geneettisen profiilin. Analyysi osoitti, että jotkin eurooppalaiset ryhmät, kuten Länsi-Saksasta, Flanderin Belgiasta, Luoteis-Ranskasta, Etelä-Ranskasta ja Espanjasta, vaikuttivat merkittävästi lähes kaikkien brittiläisten klustereiden syntymiseen. Toiset ovat vaikuttaneet vain joihinkin tiettyihin klustereihin, esimerkiksi norjalaisilla oli merkittävä vaikutus klustereiden muodostumiseen Orkneyssa ja vähemmän - Skotlannissa ja Pohjois-Irlannissa.
Geenipooli heijastaa historiaa
Geneettiset tiedot ovat heijastaneet jälkiä muuttoliikkeistä Brittein saarille viimeisen 10 tuhannen vuoden aikana. Mannerryhmien brittiklustereihin kohdistuvan vaikutuksen luonteen mukaan tutkijat erottavat muuttoliiketapahtumien suhteellisen ajan. Ne ryhmät, joiden osuus löytyy kaikista klustereista, viittaavat ilmeisesti muinaisempaan vaellukseen. Siitä on kulunut tarpeeksi aikaa, jotta heidän haplotyypit leviäisivät muualle Iso-Britanniaan. Manner jalanjälki yksittäisissä klusteissa näyttää olevan seurausta uudemmista muuttoista.

Brittein saarten asutuksen tärkeimmät tapahtumat.
Merkittävimmän jäljen jätti anglosaksien Ison-Britannian valloitus 500-luvulla. Anglosaksisen muuttoliikkeen geneettinen osuus on merkittävä Kaakkois-, Keski- ja Etelä-Englannissa, vaikka se ei ylitä 50 prosenttia missään, ja suurimmalla osalla aluetta se vaihtelee 10–40 prosenttia. Tämä osoittaa, että anglosaksit eivät täysin syrjäyttäneet brittien (kelttien) paikallista väestöä, vaan sekoittuneet siihen. Jotkut klusterit osuvat topografisesti yhteen anglosaksisten kuningaskuntien ja klaanien alueiden kanssa. Vaikuttaa siltä, että nämä alueet ovat säilyttäneet etnografisen ja geneettisen identiteettinsä vuosisatojen ajan, tutkijat korostavat.
Orkneysaarten populaatioissa 25 % geenipoolista kuuluu norjalaista alkuperää oleville esivanhemmille, nämä ovat jälkiä norjalaisten viikinkien hyökkäyksestä saarille, joka tapahtui 800-luvulla. Geneettiset tiedot osoittavat myös, että norjalaiset viikingit eivät syrjäyttäneet alkuperäisväestöä, vaan sekoittuneet sen kanssa. Mutta selkeitä geneettisiä jälkiä tanskalaisista viikingeistä, jotka hallitsivat merkittävää osaa Englannista, ei voitu löytää.
Mitä tulee Brittisaarten varhaisimpaan asutukseen - viimeisen jääkauden päätyttyä - sen jäljet ovat parhaiten säilyneet Walesin populaatioissa. Tiedot viittaavat myös myöhempään merkittävään muuttoon mantereelta, jo ennen kuin Rooman valtakunta valloitti Britannian ensimmäisen vuosituhannen alussa. Tämän muuttoliikkeen geneettinen vaikutus levisi Englantiin, Skotlantiin ja Pohjois-Irlantiin, mutta käytännössä ohitti Walesin, joka on säilyttänyt vanhimman geenipoolin.
Tutkimuksen tekijät ovat osoittaneet lopullisesti, kuinka genetiikka voi täydentää ja jalostaa arkeologista ja kielellistä tietoa populaatioiden historian rekonstruoimiseksi. Yksityiskohtaisen maan geneettisen kartan saamisessa ja brittiväestön menneisyyden tarkastelussa heitä auttoivat pätevä lähtötietojen valinta ja tehokkaat tilastolliset menetelmät.
Lähde:
Brittiväestön hienolaatuinen geneettinen rakenne
Stephen Leslie, Bruce Winney, Garrett Hellenthal, Dan Davison, Abdelhamid Boumertit, Tammy Day, Katarzyna Hutnik, Ellen C. Royrvik, Barry Cunliffe, Wellcome Trust Case Control Consortium, International Multiple Sclerosis Genetics Consortium, Daniel J. Lawson, Colin Daniel Falush Freeman, Matti Pirinen, Simon Myers, Mark Robinson, Peter Donnelly ja Walter Bodmer
Luonto, 2015, v. 519, 7543, doi: 10.1038 / nature14230
http://www.nature.com/nature/journal/v519/n7543/full/nature14230.html









