แผนที่ทางพันธุกรรมแบบโต้ตอบของโลก พันธุศาสตร์ของ Russian Ukrainians, Belarusians และ Tatars, Slavs และ Caucasians, Jews, Finns และประชากรอื่น ๆ
ในการศึกษานี้ ใช้ SNP autosomal มากกว่า 80,000 รายการ ซึ่งเป็นคะแนนที่เชื่อถือได้ ซึ่งเป็นความละเอียดที่สูงมาก ซึ่งทำให้สามารถจับอิทธิพลที่ค่อนข้างเล็กในระดับพันธุกรรมในกลุ่มคนได้ ข้อมูลการวิเคราะห์เปรียบเทียบนำมาจากการศึกษาแบบเปิดโดย V.Verenich ผู้เชี่ยวชาญด้านการวิเคราะห์เปรียบเทียบองค์ประกอบทางพันธุกรรม เครื่องคำนวณทางพันธุกรรมนั้นอยู่ในบริการ GedMatch และอนุญาตให้ทุกคนค้นหาตำแหน่งเปรียบเทียบของพวกเขาบนกราฟพันธุกรรม เมื่อต้องการทำเช่นนี้ ก็เพียงพอที่จะมีผลการทดสอบ autosomal จาก FTDNA หรือ 23andMe เมื่อสิ้นสุดการศึกษา จะมีแผนที่ของการกระจายทางภูมิศาสตร์และความถี่สูงสุดสำหรับส่วนประกอบ autosomal หลักจากโครงการ MDLP World-22
กราฟด้านล่างแสดงองค์ประกอบหลักและเปอร์เซ็นต์เฉลี่ยสำหรับประชากรแต่ละกลุ่ม ในหนึ่งบรรทัด - การแบ่งเปอร์เซ็นต์สำหรับหนึ่งประชากร แต่ละดิวิชั่น (แถบแนวตั้ง) มีค่าเท่ากับ 10% และชื่อของส่วนประกอบออโตโซมจะอยู่ในลำดับเดียวกันจากซ้ายไปขวาเช่นเดียวกับในคำอธิบายจากบนลงล่าง ยิ่งสัดส่วนร้อยละขององค์ประกอบทางพันธุกรรมร่วมกันระหว่างชนชาติต่างๆ ใกล้เคียงกันมากเท่าใด ตัวเลขในกราฟด้านบนก็จะยิ่งคล้ายกันมากขึ้นเท่านั้น มาเริ่มกันเลย...
พันธุศาสตร์ของชาวเยอรมัน ลิทัวเนีย รัสเซีย สวีเดน ฟินน์ ฯลฯ
กราฟนี้แสดงองค์ประกอบทางพันธุกรรมหลักของชนชาติยุโรปและสอดคล้องกับการลดลงขององค์ประกอบยุโรปตะวันออก (ยุโรปตะวันออกเฉียงเหนือ) ในประชากรต่างๆ ดังที่คุณเห็นแล้ว ชนชาติยุโรปทั้งหมดมีความแตกต่างกันในแง่พันธุกรรม และมีองค์ประกอบทางพันธุกรรมในชุดต้นกำเนิดเดียวกัน กระนั้นก็ตามในเปอร์เซ็นต์ที่แตกต่างกันมาก สำหรับชาวสลาฟและบอลต์ทั้งหมด โดยทั่วไปแล้ว หนึ่งในองค์ประกอบที่สำคัญที่สุดคือองค์ประกอบของยุโรปตะวันออก ซึ่งอยู่สูงสุดในหมู่ชาวลิทัวเนียและเบลารุส น่าจะเป็นตั้งแต่สมัย "วัฒนธรรมเครื่องมีสาย" ทางโบราณคดีอาณาเขตของประเทศเหล่านี้เป็นศูนย์กลางของแหล่งกำเนิดขององค์ประกอบนี้ มีตัวแทนมากกว่า 80% ในหมู่ชาวลิทัวเนียและมีเพียง 20% ในหมู่ชาวอิตาลีสีม่วงแสดงถึงองค์ประกอบแอตแลนโต-เมดิเตอร์เรเนียน และจะเพิ่มขึ้นเมื่อเคลื่อนจากตะวันออกเฉียงเหนือไปตะวันตกเฉียงใต้ ดังนั้นในกลุ่ม Finns จะมีค่าเฉลี่ยถึง 15% และในหมู่ชาวอิตาลี 40% ส่วนประกอบที่เหลือมีความเด่นชัดน้อยกว่า
พันธุศาสตร์ของ Russian Ukrainians Belarusians

กราฟนี้แสดงให้เห็นถึงชาวสลาฟตะวันออก - รัสเซีย เบลารุส ยูเครน. ความสนใจถูกดึงดูดไปยังความคล้ายคลึงของรูปแบบทางพันธุกรรมของชนชาติทั้งสามที่อยู่ในรายการ และภายในขอบของข้อผิดพลาด พวกมันแตกต่างกันเล็กน้อยมาก - ยูเครนและรัสเซียตอนใต้มีการเพิ่มขึ้นเล็กน้อยในองค์ประกอบเอเชียตะวันตก และรัสเซียตอนเหนือเพิ่มขึ้นเล็กน้อยในหนึ่ง ขององค์ประกอบไซบีเรียซึ่งมีเงื่อนไขเรียกว่า Samoyedic และส่วนประกอบที่เพิ่มขึ้นของ Mesolithic of Europe เป็นประมาณ 10% ซึ่งตามตัวบ่งชี้หลังทำให้พวกเขาใกล้ชิดกับประชากรสแกนดิเนเวียที่พูดภาษาเยอรมัน - ชาวสวีเดนมากขึ้น
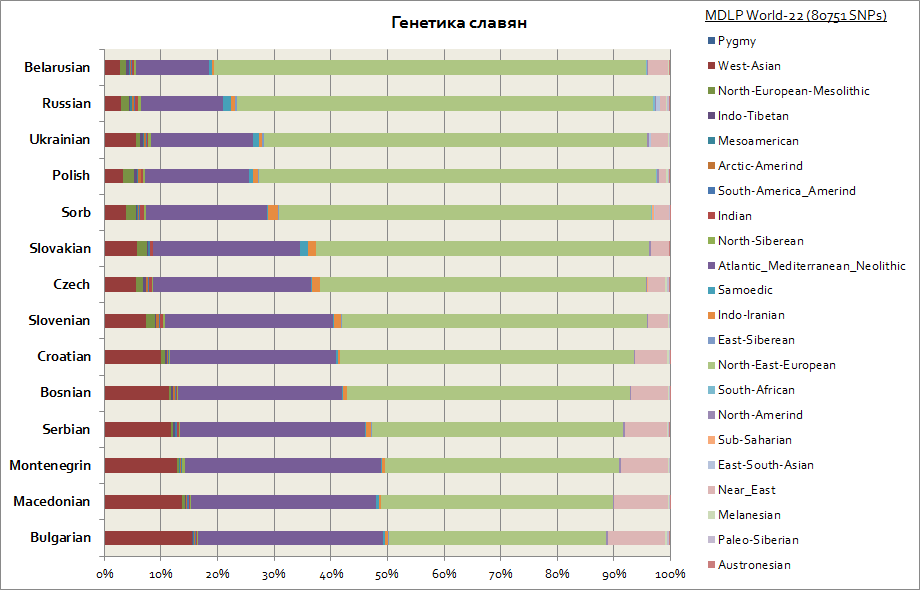
แผนภูมินี้แสดงภาพชาวสลาฟทั้งหมด รวมทั้งชาวตะวันตก - โปแลนด์และเช็ก เช่นเดียวกับทางใต้ - เซิร์บ บัลแกเรีย มาซิโดเนีย เป็นต้น
ชาวสลาฟทั้งหมดมี 2 องค์ประกอบหลัก ได้แก่ ยุโรปตะวันออกและแอตแลนโต-เมดิเตอร์เรเนียน อย่างแรกคือค่าสูงสุดสำหรับชาวเบลารุส และครั้งที่สองสำหรับชาวสลาฟใต้ทั้งหมด - เซิร์บ มาซิโดเนีย และบัลแกเรีย องค์ประกอบของยุโรปตะวันออกนั้นมีต้นกำเนิดมาจากชาวสลาฟมากกว่า และองค์ประกอบแอตแลนโต-เมดิเตอร์เรเนียนนั้นได้มามากกว่าเมื่อชาวสลาฟอพยพไปยังคาบสมุทรบอลข่าน ชาวยูเครนตะวันตกและสโลวักมีการเพิ่มขึ้นเล็กน้อยในองค์ประกอบ Samoyedic เมื่อเทียบกับชนชาติสลาฟที่อยู่ใกล้เคียง - เบลารุส, เช็ก, โปแลนด์; นี่อาจเป็นร่องรอยทางพันธุกรรมของการอพยพในยุคกลางของชาวฮั่นและชาวอูเกรียนไปยังยุโรปกลาง
พันธุศาสตร์ของชาวสลาฟ รัสเซีย และตาตาร์ เยอรมัน คอเคเซียน ยิว ฯลฯ
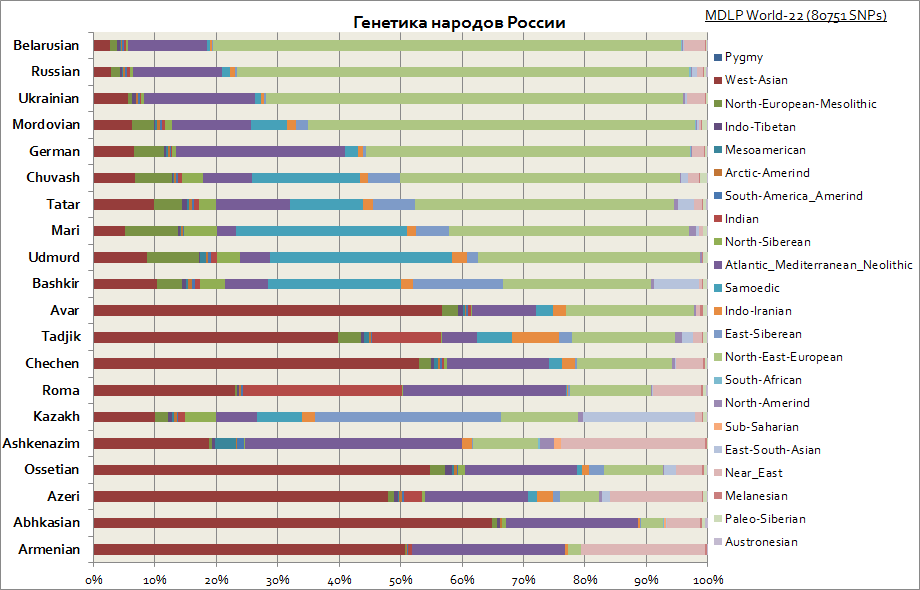
กราฟนี้แสดงต้นกำเนิดต่างๆ ของชนชาติรัสเซีย ดังที่เห็นได้ในหมู่ชาวสลาฟองค์ประกอบหลักคือองค์ประกอบหลักของยุโรปตะวันออกในขณะที่ผู้คนในภูมิภาคโวลก้าส่วนแบ่งขององค์ประกอบไซบีเรียเพิ่มขึ้น ในขณะที่สำหรับคอเคเซียน องค์ประกอบในเอเชียตะวันตก ทะเลเมดิเตอร์เรเนียน และตะวันออกกลางมีลักษณะเฉพาะมากที่สุด
พันธุศาสตร์ของ Finns, Ugrians, Udmurts, Hungarians, Saami เป็นต้น

อย่างที่เห็น Finns, Veps และ Karelians มีต้นกำเนิดทางพันธุกรรมที่คล้ายคลึงกันกับ Slavs พวกเขายังมีองค์ประกอบที่ใหญ่ที่สุดของยุโรปตะวันออกโดยลดลงใกล้กับเทือกเขาอูราลและภูมิภาคโวลก้าด้วยการเพิ่มขึ้นของส่วนประกอบไซบีเรียในภูมิภาคนี้ นอกจากนี้ ชนชาติ Finno-Ugric ทั้งหมดยังมีองค์ประกอบสำคัญของ Mesolithic of Europe ซึ่งเข้าถึงเกือบ 80% ในหมู่ชาวซามิและเกี่ยวข้องกับประชากรก่อนอินโด - ยูโรเปียนและก่อนยุคหินใหม่ของยุโรป สำหรับชาวฮังกาเรียนโดยรวม ชุดขององค์ประกอบทางพันธุกรรมเหล่านั้นมีลักษณะเฉพาะ เช่นเดียวกับประชากรอื่นๆ ของภูมิภาคคาร์เพเทียนและยุโรปกลาง
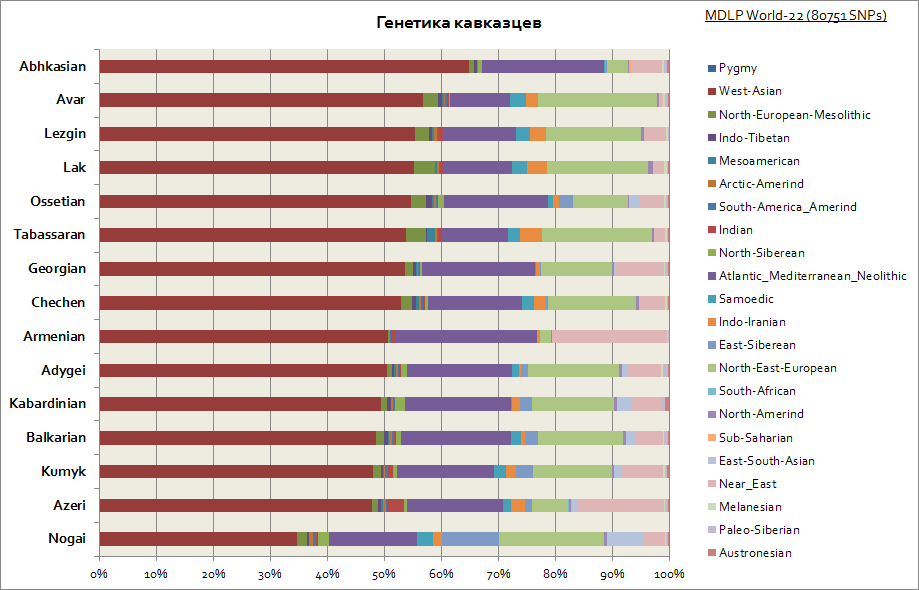
ดังที่เห็นได้ชัดเจน คอเคซัสทั้งหมดมีลักษณะเฉพาะด้วยแหล่งกำเนิดทางพันธุกรรมที่ค่อนข้างคล้ายคลึงกัน ซึ่งเป็นสัดส่วนที่มากขององค์ประกอบในเอเชียตะวันตกและทะเลเมดิเตอร์เรเนียน มีเพียง Nogais เท่านั้นที่โดดเด่นเพียงเล็กน้อย - พวกเขามีส่วนแบ่งส่วนประกอบไซบีเรียเพิ่มขึ้น

ดังที่เห็นได้ในอาซเกนาซิมและเซฟาร์ดิม มีความถี่สูงของส่วนประกอบเอเชียตะวันตก แอตแลนโต-เมดิเตอร์เรเนียน และตะวันออกกลาง ในเวลาเดียวกัน Ashkenazim มีการเพิ่มขึ้นเล็กน้อยในองค์ประกอบไซบีเรียซึ่งอาจเป็นเพราะมรดก Khazar และเพิ่มขึ้นถึง 30% ขององค์ประกอบยุโรปตะวันออกซึ่งตามตัวบ่งชี้นี้ทำให้พวกเขาใกล้ชิดกับ ประเทศทางตอนใต้ของยุโรป
เฉพาะชาวยิวเอธิโอเปียและชาวยิวอินเดียเท่านั้นที่ถูกคัดออกจาก "บริษัท" ของพวกเขาโดยเฉพาะ แอฟริกาเหนือมีสัดส่วนที่สูงของแอฟริกาตอนใต้สะฮารา (มากถึง 40%) ในขณะที่หลังมีส่วนแบ่งขององค์ประกอบทางพันธุกรรมของเอเชียใต้ เรียกว่าอินเดียอย่างมีเงื่อนไข (มากถึง 50%)
พันธุศาสตร์ของ Tatars, Bashkirs, Azerbaijanis, Chuvashs เป็นต้น
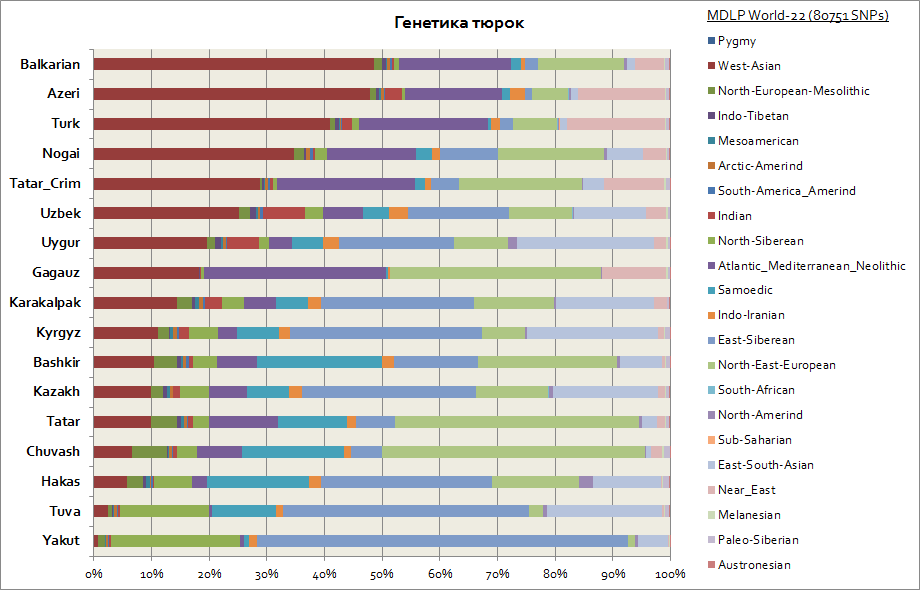
ชาวเติร์กในแง่พันธุกรรมกลายเป็นกลุ่มชาติพันธุ์ที่ต่างกันมากที่สุดกลุ่มหนึ่ง เนื่องจากองค์ประกอบทางพันธุกรรมของพวกมันแตกต่างกันอย่างมาก ดังนั้น เนื่องจากบ้านเกิดหลักของพวกเติร์กคือไซบีเรีย ดังนั้นผู้คนเช่น Yakuts, Tuvans, Khakasses จึงยังคงรักษาองค์ประกอบ autosomal ของไซบีเรียตะวันออกไว้เป็นเปอร์เซ็นต์ที่ใหญ่ที่สุดซึ่งถึง 30 ถึง 65% ในพวกเขา องค์ประกอบทางพันธุกรรมนี้ยังเป็นองค์ประกอบหลักในหมู่คีร์กีซและคาซัคสถาน ส่วนประกอบที่เหลือทำให้ชาวเติร์กใกล้ชิดกับประชาชนจากภูมิภาคที่พำนักมากขึ้น ดังนั้นสำหรับยาคุตและทูแวน สิ่งเหล่านี้คือส่วนประกอบทางเหนือของไซบีเรียและซามอยด์ โดยรวมแล้ว 3 องค์ประกอบของไซบีเรียนในบรรดายาคุตนั้นคิดเป็น 90% ในบรรดาชาวทูแวนมากถึง 70% โดยเพิ่มขึ้นเป็น 20% ขององค์ประกอบเอเชียตะวันออก - ใต้ ซึ่งสัมพันธ์กับกระแสการอพยพของประชากรในเอเชียตะวันออกในระดับที่มากขึ้น สำหรับ Bashkirs ส่วนแบ่งขององค์ประกอบ 3 ไซบีเรียสูงถึง 45% และส่วนประกอบในเอเชียตะวันออกเฉียงใต้สูงถึง 10% ตาตาร์มีข้อมูลขององค์ประกอบทางพันธุกรรมของไซบีเรีย 3 โดยเฉลี่ยจาก 25 ถึง 50% ในเวลาเดียวกันสัดส่วนของส่วนประกอบที่เป็นลักษณะเฉพาะของประชากรคอเคซอยด์ในบัชคีร์นั้นสูงถึง 45% และโดยเฉลี่ยแล้วในหมู่พวกตาตาร์จาก 50 ถึง 70% พันธุศาสตร์ของอาเซอร์ไบจานและเติร์กแทบไม่แตกต่างกันภายในขอบของข้อผิดพลาด พวกเขาเช่นเดียวกับชนชาติอื่น ๆ ของคอเคซัสและทรานคอเคเซียมีองค์ประกอบที่สำคัญของเอเชียตะวันตก (ถึง 50%) และองค์ประกอบแอตแลนโต - เมดิเตอร์เรเนียน (มากถึง โดยเฉลี่ย 20%) ส่วนแบ่งขององค์ประกอบไซบีเรีย 3 ตัวแสดงโดยอาเซอร์ไบจาน เติร์กและบัลการ์ - ที่ระดับ 3-7%
บทสรุป
พันธุกรรมของประชาชนไม่มีความสัมพันธ์โดยตรงและสำคัญกับการกระจายตัวของตระกูลภาษา หรือร้อยละของเครื่องหมายที่มีผู้ปกครองเดียว - Y-DNA และ mt-DNA haplogroups ที่แสดงในประชากรเฉพาะ ความสัมพันธ์ที่ยิ่งใหญ่ที่สุดสามารถสืบย้อนได้ตามหลักการของอาณาเขต-ภูมิศาสตร์ ดังนั้นสัดส่วนขององค์ประกอบไซบีเรียซึ่งโดยทั่วไปมีลักษณะเฉพาะของเผ่าพันธุ์มองโกลอยด์จึงค่อยๆ ลดลงจากตะวันออกไปตะวันตก และสัดส่วนของลักษณะส่วนประกอบของเผ่าพันธุ์คอเคเซียนก็เพิ่มขึ้นตามลำดับ ในดินแดนชายแดนตามแนวเหนือของเทือกเขาอูราลถึงเอเชียกลางอัตราส่วนจะเท่ากันโดยประมาณ ในภูมิภาคทางตะวันออกของทะเลสาบไบคาลลักษณะองค์ประกอบทางพันธุกรรมของเผ่าพันธุ์คอเคเซียนขนาดใหญ่นั้นแทบจะไม่มีตัวแทนในขณะเดียวกันในภูมิภาคทางตะวันตกของสาย Pechora-Volga ลักษณะขององค์ประกอบไซบีเรียของเผ่าพันธุ์มองโกลอยด์ขนาดใหญ่จะหายไป .การแพร่กระจายขององค์ประกอบทางพันธุกรรมของยุโรปตะวันออกไปยังไซบีเรียเกิดขึ้นอย่างมากในยุคสำริด (วัฒนธรรมของ Andronov Circle) แม้ว่ายอดเขาส่วนบุคคลทางตะวันออกสุดของไซบีเรียท่ามกลาง Chukchi อาจเกี่ยวข้องกับการอพยพของรัสเซียใน ศตวรรษที่ 17.
ส่วนแบ่งขององค์ประกอบ Sub-Saharan ซึ่งเป็นลักษณะของเผ่าพันธุ์ Negroid นั้นกระจายไปทั่วแอฟริกา - จนถึงทะเลเมดิเตอร์เรเนียนตอนใต้และชายแดนทางเหนือของทวีปแอฟริกาถึงจุดสูงสุดในส่วนเส้นศูนย์สูตรและแทบไม่เคยพบนอกมันเลย พื้นหลังสีอ่อนกระจายอยู่บนคาบสมุทรอาหรับและทางตอนใต้ของที่ราบสูงอิหร่าน
ภูมิศาสตร์ขององค์ประกอบทางพันธุกรรม
อเล็กซี่ ซอร์ริน
โครงการ
การวิเคราะห์ทางพันธุศาสตร์ได้เข้าสู่ขั้นตอนการปฏิบัติงานตามปกติมาอย่างยาวนานและมั่นคงในด้านความรู้ประยุกต์ เช่น นิติวิทยาศาสตร์และการแพทย์ การปรับปรุงวิธีการรับรู้เครื่องหมายทางพันธุกรรมพบว่ามีการประยุกต์ใช้ในปัญหาทางวิทยาศาสตร์ขั้นพื้นฐาน
ตัวอย่างเช่น ขอบคุณโซลูชันทางเทคนิคใหม่ mtDNA . ที่จัดลำดับอย่างสมบูรณ์มนุษย์ยุคก่อนประวัติศาสตร์ในประเภท Neanderthal และในอนาคตอันใกล้นี้ นักวิทยาศาสตร์หวังว่าจะสามารถถอดรหัสจีโนมนิวเคลียร์ที่สมบูรณ์และเปรียบเทียบกับมนุษย์ได้ ประวัติศาสตร์ของเราถูกเปิดเผยแก่เรามากขึ้นเรื่อยๆ ไม่ใช่ผ่านการค้นพบทางโบราณคดี แต่ผ่านวิธีการวิเคราะห์ทางพันธุกรรม ตัวอย่างเช่น เราเรียนรู้ลำดับเหตุการณ์โดยประมาณของการตั้งถิ่นฐานของโลกและพบว่าในช่วงเริ่มต้นของการพัฒนา มนุษยชาติเกือบ หายไปจากพื้นโลกเพราะภัยแล้ง
อย่างไรก็ตาม ข้อมูลที่เข้ารหัสในนิวคลีโอไทด์หลายพันล้านตัวของ DNA สามารถบอกข้อมูลเพิ่มเติมเกี่ยวกับบุคคลหนึ่งๆ และเกี่ยวกับมนุษยชาติโดยรวมได้มาก John Novembre แห่งมหาวิทยาลัยแคลิฟอร์เนียแห่งลอสแองเจลิสและเพื่อนร่วมงานของเขาแสดงให้เห็นว่าการวิเคราะห์ทางพันธุกรรมขนาดใหญ่ของชาวยุโรปมากกว่าสามพันคนทำให้ทุกวันนี้สามารถเปิดเผยโครงสร้างของประชากรยุโรปและค้นหาว่าตัวแทนที่แตกต่างกันทางพันธุกรรมของชาวยุโรปขนาดเล็กได้อย่างไร รัฐคือ
ตอนนี้นักวิทยาศาสตร์สามารถกำหนดสถานที่กำเนิดของชาวยุโรปที่มีความแม่นยำหลายร้อยกิโลเมตรโดยอาศัย DNA ของเขาเท่านั้น
มีวิธีทางคณิตศาสตร์มากมายในการแก้ปัญหาที่ไม่ชัดเจนนัก กรรมพันธุ์ที่พบมากที่สุดคือวิธีการขององค์ประกอบหลัก ซึ่งช่วยให้คุณกำหนด "แกน" ดังกล่าวในอาร์เรย์ข้อมูลหลายมิติ ซึ่งจะเห็นความแตกต่างของจีโนมได้ชัดเจนที่สุด Novembre และเพื่อนร่วมงานของเขาใช้สิ่งเหล่านี้โดย จำกัด ตัวเองให้เหลือสองแกน
เมื่อวางจุดเกือบหนึ่งพันห้าพันจุดไว้บนเครื่องบินแล้ว พวกเขาเห็นภาพที่ชวนให้นึกถึงแผนที่การเมืองของยุโรปอย่างน่าทึ่ง
จีโนมของโปรตุเกสและสเปนอยู่บนแผนที่นี้ทางด้านซ้ายของฝรั่งเศส ชาวเยอรมันอยู่ทางขวา "ชาวอิตาลี" อยู่ใต้ฝรั่งเศส และอังกฤษอยู่เหนือพวกเขา โชคดีที่คุณสามารถหมุนแผนที่ผลลัพธ์ได้ตามต้องการ ซึ่งจะไม่เปลี่ยนความสัมพันธ์ทางเรขาคณิตระหว่างตำแหน่งของจุดต่างๆ สิ่งเดียวที่เหลือให้นักวิทยาศาสตร์ทำคือเปรียบเทียบพล็อตทางพันธุกรรมกับแผนที่ทางภูมิศาสตร์ - ที่เรียกว่า clinality ตามพารามิเตอร์คอมโพสิตที่สอดคล้องกับแกนหลัก - และพิจารณาว่า "องค์ประกอบหลัก" ที่สำคัญที่สุดสอดคล้องกับทิศใต้ -ตะวันออกเฉียงใต้ - แกนเหนือ - ตะวันตกเฉียงเหนือและแม่นยำในแบบฟอร์มนี้และนำเสนอแผนที่ให้เพื่อนร่วมงาน
"แผนที่พันธุกรรมของยุโรป" โดย John Novembre และเพื่อนร่วมงาน ที่ด้านล่างขวาเป็นผลงานที่คล้ายคลึงกันโดย Manfred Kaiser ซึ่งเผยแพร่เมื่อเดือนที่แล้ว // John Novembre/M.Kaiser
แน่นอนว่าแผนที่สุดท้ายไม่มีข้อบกพร่อง จีโนมส่วนบุคคลไม่เพียงแต่แผ่ขยายออกไปไกลเกินขอบเขตของประเทศที่สอดคล้องกับพวกมันเท่านั้น รัสเซียตั้งอยู่บนแผนภูมินี้ท่ามกลางชาวเช็กและชาวโปแลนด์ โดยอยู่ "ตะวันตก" ของชาวยูเครน และโดยทั่วไปแล้วชาวสโลวักจะตกอยู่ใต้เทือกเขาแอลป์ ซึ่งปรากฏขึ้นที่ปลายสุดของ "รองเท้าบูท" ของอิตาลี อย่างไรก็ตาม สิ่งนี้สามารถอธิบายได้ด้วยลักษณะเฉพาะของตัวอย่างที่น้อยของทั้งสอง: มีชาวรัสเซียเพียงหกคนในข้อมูลเริ่มต้น และพบสโลวักเพียงคนเดียว
อย่างไรก็ตาม ด้วยการใช้แผนที่นี้เท่านั้น บ้านเกิดของผู้คน 90% สามารถระบุได้อย่างแม่นยำ 700 กม. และ 50% - สูงสุด 300 กม.
แม้ว่า DNA จะมี "ตัวอักษร" 200,000 ตัวน้อยกว่า 0.01% ของรหัสพันธุกรรมมนุษย์ทั้งหมด
กราฟยังแสดงให้เห็นอย่างชัดเจนว่าความหลากหลายทางพันธุกรรมของชาวยุโรปตอนใต้มีความสำคัญมากกว่าคนทางตอนเหนือมาก ข้อมูลนี้ ตามโนเวมเบรเองซึ่งสามารถอธิบายได้อย่างง่ายดายจากประวัติศาสตร์การตั้งถิ่นฐานของมนุษย์ในยุโรป หลังจากการปรากฏตัวของ Homo sapiens ที่นี่เมื่อ 35,000 ปีก่อน เพียง 15,000 ปีต่อมา ด้วยการถอยของธารน้ำแข็ง การอพยพจำนวนมากครั้งแรกของผู้คนไปทางเหนือก็เกิดขึ้น และหลังจากนั้นอีก 10,000 ปี กับการถือกำเนิดของเกษตรกรรม ชาวใต้กลุ่มใหม่เดินทางไปทางเหนือ ซึ่งนับพันปีเหล่านี้เป็นตัวแทนของประชากรที่มีรูปแบบดีน่าประทับใจ
เหมือนกัน งานโดยอาศัยข้อมูลทางพันธุกรรมเดียวกันเป็นส่วนใหญ่ ได้รับการตีพิมพ์ในวารสาร Current Biology เมื่อเดือนที่แล้ว เป็นที่น่าสังเกตว่าผู้นำ Manfred Kaiser ยังสามารถได้รับภาพที่คล้ายคลึงกันแม้ว่าจะไม่ฉ่ำมากนักโดยใช้วิธีการองค์ประกอบหลักเดียวกัน การวิเคราะห์นั้นเกี่ยวข้องกับ 2,500 จีโนมที่รวบรวมอย่างระมัดระวังโดย GlaxoSmithKline บริษัทที่ค้นหายีนที่รับผิดชอบต่อผลข้างเคียงของผู้ที่ใช้ยาหลายชนิด

"แผนที่ทางพันธุกรรม" ของประชากรสวิตเซอร์แลนด์ // John Novembre
ในเวลาเดียวกัน วิธีการขององค์ประกอบหลักในทั้งสองกรณีล้มเหลวในการแยกชาวยุโรปออกเป็นกลุ่มทางพันธุกรรมที่แยกจากกัน กล่าวอีกนัยหนึ่ง ความหลากหลายทางพันธุกรรมของชาวยุโรปมีขนาดเล็กมากจนแม้แต่ mononucleotide polymorphisms ครึ่งล้านก็ยังไม่เพียงพอที่จะแบ่งคนเพียงสามพันคนออกเป็นกลุ่มย่อยเล็กๆ
“บางครั้งเราชอบพูดถึงยีน “รัสเซียน” “ยีนโปแลนด์” หรือ “ตาตาร์” แต่บทความนี้แสดงให้เราเห็นชัดเจนว่าไม่มียีนดังกล่าว!”
- Oleg Balanovsky หนึ่งในผู้เชี่ยวชาญหลักในประเทศในด้านภูมิศาสตร์ยีนจากห้องปฏิบัติการพันธุศาสตร์ประชากรมนุษย์ของศูนย์วิจัยพันธุศาสตร์การแพทย์ของ Russian Academy of Medical Sciences ให้ความเห็นเกี่ยวกับงานเหล่านี้กับ Gazeta.Ru เป็นที่รู้จักกันดีถึงผู้อ่านของเรา
“ด้วยการศึกษายีนหนึ่งแสนยีนเท่านั้น ผู้เขียนสามารถ (และถึงแม้จะผิดพลาดหลายร้อยกิโลเมตร) เพื่อระบุแหล่งกำเนิดที่เป็นไปได้ของชาวยุโรปที่ศึกษา เป็นไปไม่ได้ที่จะทำเช่นนี้กับยีนหนึ่งยีนและร้อยยีน! บาลานอฟสกีกล่าว - ไม่มีลักษณะเฉพาะของยีนเดียวของประชากรในประเทศที่กำหนด มีเพียงการรวมกันของยีนหลายแสนตัวเท่านั้นที่มีความเฉพาะเจาะจงไม่มากก็น้อย
นี่หมายความว่าข้อโต้แย้งของบรรดาผู้เกรงกลัว ส่งผลกระทบต่อยีนพูลของประชาชนผ่านยีนเฉพาะของมัน"
“เรารอบทความนี้มานานแล้ว เทคโนโลยีในการวิเคราะห์เครื่องหมายทางพันธุกรรมนับล้านมีมาหลายปีแล้ว และในบทความนี้ นักวิทยาศาสตร์ได้นำเทคโนโลยีใหม่เหล่านี้มาใช้เพื่อศึกษากลุ่มยีนของยุโรปในที่สุด – งานนี้แสดงให้เห็นถึงความต่อเนื่องทางวิทยาศาสตร์ได้อย่างสมบูรณ์แบบ: เมื่อ 10-15 ปีที่แล้วมีงานเกี่ยวกับ DNA ของยล (สายการสืบทอดของมารดา) และโครโมโซม Y (สายการสืบทอดของบิดา) การพัฒนาครั้งแรกก็เกิดขึ้นเช่นกัน กลุ่มยีนของยุโรปและแม้แต่ข้อสรุปเกี่ยวกับความสำคัญทางภูมิศาสตร์ในการจัดลำดับกลุ่มยีนก็เหมือนกับตอนนี้
จากข้อมูลของ Balanovsky เทคโนโลยีในปัจจุบันช่วยให้ได้ผลลัพธ์ที่น่าเชื่อถือและแม่นยำอย่างที่ไม่เคยมีมาก่อน: “สิ่งที่พันธุศาสตร์แทบไม่มองเห็นได้ในหมอกหนาทึบเมื่อสี่สิบปีที่แล้ว สิ่งที่เมื่อสิบปีก่อนเริ่มส่องสว่างด้วยไฟตัดหมอกอันทรงพลังสองดวง (ดีเอ็นเอของไมโตคอนเดรียและโครโมโซม Y) เรา ตอนนี้เริ่มมองเห็นสิ่งเดียวกันในแง่ของแสงที่อ่อนแอกว่าล้านแสง (ยีนถึงแม้จะให้ข้อมูลน้อยกว่า แต่มีจำนวนมากและกระจายไปทั่วจีโนม)
“ขั้นตอนต่อไปและขั้นตอนสุดท้าย - การส่องสว่างเต็ม 100% - สามารถคาดหวังได้ในอีกไม่กี่ปีข้างหน้า เมื่อมีการจัดลำดับจีโนมทั้งหมดอย่างเต็มรูปแบบ - ไม่ใช่นิวคลีโอไทด์หนึ่งล้านอย่างที่เป็นอยู่ตอนนี้ แต่เป็นพันล้านที่มีอยู่ในจีโนม ” ผู้เชี่ยวชาญในประเทศคาดการณ์
ในความเห็นของเขา คำถามเดียวที่เกิดขึ้นคือ จำเป็นต้องรอข้อมูลรายละเอียดดังกล่าวหรือไม่ อันที่จริงแล้ว "แสงอันทรงพลังทั้งสองของเรา (ดีเอ็นเอของไมโตคอนเดรียและโครโมโซม Y) และแสงที่อ่อนแอกว่าล้านดวง (แต่กระจายไปทั่วจีโนม) ทำให้เราเห็นโครงสร้างของกลุ่มยีนของมนุษย์และวาดทั้งทางวิทยาศาสตร์ขั้นพื้นฐานและประยุกต์ ข้อสรุป”
นักวิทยาศาสตร์กังวลเกี่ยวกับความจริงที่ว่าจนถึงขณะนี้จำนวนของยีนที่ศึกษาในจีโนมนั้นมากกว่าจำนวนคนที่ได้รับการศึกษายีนเหล่านี้มาก
"เฉพาะการศึกษากลุ่มยีน - ผลรวมของยีน - ทำให้เราสามารถศึกษาพันธุศาสตร์ของผู้คนเพื่อช่วยให้แพทย์วินิจฉัยและทุกคน - เพื่อติดตามประวัติของบุคคลหรือครอบครัวของพวกเขาเอง" สรุป โอเล็ก บาลานอฟสกี
5 มกราคม 2556พิมพ์
เราได้ยินมาโดยตลอดว่าชาวรัสเซียไม่ใช่คนเลือดแข็ง มีความสัมพันธ์ทางสายเลือด แต่เป็นกลุ่มคนที่รวมกันเป็นหนึ่งด้วยวัฒนธรรมและอาณาเขตร่วมกัน ทุกคนจำวลีเด็ดของปูตินที่ว่า "ไม่มีชาวรัสเซียบริสุทธิ์!" และ "เการัสเซียทุกคนคุณจะพบตาตาร์อย่างแน่นอน"
สมมติว่าเรา "ต่างกันมากในเลือด" "ไม่ได้งอกมาจากรากเดียวกัน" แต่เป็นหม้อหลอมละลายของพวกตาตาร์ คอเคเซียน เยอรมัน ฟินแลนด์ บูรยัต มอร์โดเวียน และชนชาติอื่นๆ ที่เคยวิ่ง เข้ามา หลงทาง ที่ดินเราก็รับหมด ให้เข้าบ้าน เลี้ยงไว้เป็นญาติกัน
เกือบจะกลายเป็นสัจธรรมที่นักการเมืองใช้ซึ่งทำให้แนวคิดของรัสเซียไม่ชัดเจนและในเวลาเดียวกันสำหรับทุกคนก็เป็นตั๋วเข้าสู่สิ่งแวดล้อมของชาวรัสเซีย
วิธีการนี้ ซึ่งได้รับการยกธงโดยองค์กร Russophobic a la la "สิทธิมนุษยชน" จำนวนมากและสื่อของรัสเซีย Russophobic ได้ท่วมท้นคลื่นวิทยุ แต่ไม่ช้าก็เร็ว ปูตินและคนอื่นๆ แบบเขายังคงต้องตอบคำกล่าวที่เหยียดหยามคนรัสเซีย คำตัดสินของนักวิทยาศาสตร์นั้นไร้ความปราณี:
1) ในปี 2552 เสร็จสิ้น "การอ่าน" (การจัดลำดับ) ของจีโนมของตัวแทนกลุ่มชาติพันธุ์รัสเซียเสร็จสมบูรณ์ นั่นคือลำดับของนิวคลีโอไทด์ทั้งหกพันล้านในจีโนมของชายรัสเซียได้รับการพิจารณาแล้ว เศรษฐกิจทางพันธุกรรมทั้งหมดของเขาอยู่ในมุมมองที่สมบูรณ์
(จีโนมมนุษย์ประกอบด้วยโครโมโซม 23 คู่: 23 คู่จากแม่, 23 คู่จากพ่อ โครโมโซมแต่ละโครโมโซมประกอบด้วยโมเลกุล DNA หนึ่งตัวที่เกิดขึ้นจากสายโซ่ของนิวคลีโอไทด์ 50-250 ล้าน จีโนมของชายรัสเซียถูกจัดลำดับจีโนมรัสเซีย ถูกถอดรหัสบนพื้นฐานของศูนย์วิจัยแห่งชาติ "สถาบัน Kurchatov" ตามความคิดริเริ่มของสมาชิกที่สอดคล้องกันของ Russian Academy of Sciences ผู้อำนวยการศูนย์วิจัยแห่งชาติ "สถาบัน Kurchatov" Mikhail Kovalchuk ตามข้อมูลที่ได้รับจาก Russian Academy of วิทยาศาสตร์ สถาบัน Kurchatov ใช้เงินประมาณ 20 ล้านดอลลาร์ในการซื้ออุปกรณ์จัดลำดับเพียงอย่างเดียว ศูนย์ "สถาบัน Kurchatov" มีสถานะทางวิทยาศาสตร์ที่เป็นที่ยอมรับในโลก)
เป็นที่ทราบกันดีว่านี่เป็นยีนถอดรหัสตัวที่เจ็ดหลังสันเขาอูราล: ก่อนหน้านั้นมียาคุต, บูรัต, จีน, คาซัค, ผู้เชื่อเก่า, คันตี นั่นคือข้อกำหนดเบื้องต้นทั้งหมดสำหรับแผนที่ชาติพันธุ์แรกของรัสเซียได้ถูกสร้างขึ้น แต่ทั้งหมดนี้เป็นจีโนมคอมโพสิต กล่าวคือ ชิ้นส่วนที่ประกอบขึ้นหลังจากถอดรหัสสารพันธุกรรมของตัวแทนที่แตกต่างกันของประชากรกลุ่มเดียวกัน
ภาพเหมือนทางพันธุกรรมเต็มรูปแบบของชายรัสเซียคนหนึ่งมีเพียงแปดในโลกเท่านั้น ตอนนี้มีคนที่จะเปรียบเทียบรัสเซียกับ: กับชาวอเมริกัน, แอฟริกัน, เกาหลี, ยุโรป ...
« เราไม่พบการแนะนำตาตาร์ที่เห็นได้ชัดเจนในจีโนมรัสเซียซึ่งหักล้างทฤษฎีเกี่ยวกับอิทธิพลการทำลายล้างของแอกมองโกล, - เน้นหัวหน้าทิศทางจีโนมที่ศูนย์วิจัยแห่งชาติ "สถาบัน Kurchatov" นักวิชาการ Konstantin Skryabin - ไซบีเรียนมีพันธุกรรมเหมือนกับผู้เชื่อในสมัยโบราณ พวกเขามีจีโนมรัสเซียเพียงตัวเดียว ไม่มีความแตกต่างระหว่างจีโนมของรัสเซียและยูเครน - หนึ่งจีโนม. ความแตกต่างของเรากับชาวโปแลนด์มีน้อย”
นักวิชาการ Konstantin Skryabin เชื่อว่า "ภายในห้าหรือหกปี แผนที่ทางพันธุกรรมของผู้คนทั้งหมดในโลกจะถูกวาดขึ้น ซึ่งเป็นขั้นตอนที่สำคัญในการทำความเข้าใจความอ่อนแอของกลุ่มชาติพันธุ์ใดๆ ต่อยา โรค และผลิตภัณฑ์" รู้สึกได้ถึงค่าใช้จ่าย... ชาวอเมริกันในทศวรรษ 1990 ให้การประมาณการดังต่อไปนี้: ค่าใช้จ่ายในการจัดลำดับนิวคลีโอไทด์หนึ่งตัวคือ $1; ตามแหล่งอื่น - มากถึง 3-5 ดอลลาร์
(การอ่านตามตัวอักษรของรหัสพันธุกรรม) ของ mitochondrial DNA และ DNA ของโครโมโซม Y ของมนุษย์เป็นวิธีการวิเคราะห์ DNA ที่ก้าวหน้าที่สุดจนถึงปัจจุบัน DNA ของไมโตคอนเดรียถูกส่งผ่านสายเพศหญิงจากรุ่นสู่รุ่นแทบไม่เปลี่ยนแปลงตั้งแต่เมื่อครั้ง "บรรพบุรุษของมนุษยชาติอีฟ" ปีนขึ้นจากต้นไม้ในแอฟริกาตะวันออก และโครโมโซม Y พบได้เฉพาะในผู้ชายและดังนั้นจึงส่งไปยังลูกหลานชายแทบไม่เปลี่ยนแปลงในขณะที่โครโมโซมอื่น ๆ ทั้งหมดเมื่อถ่ายทอดจากพ่อและแม่สู่ลูก ถูกสับเปลี่ยนโดยธรรมชาติเหมือนสำรับไพ่ก่อนแจกจ่าย ดังนั้น ไม่เหมือนสัญญาณทางอ้อม (ลักษณะที่ปรากฏ สัดส่วนของร่างกาย) การจัดลำดับของ DNA ยลและ DNA ของโครโมโซม Y อย่างไม่อาจโต้แย้งได้และบ่งบอกถึงระดับความสัมพันธ์ของบุคคลโดยตรง)
2) นักมานุษยวิทยาดีเด่น นักวิจัยธรรมชาติชีวภาพของมนุษย์ A.P. Bogdanov เขียนเมื่อปลายศตวรรษที่ 19 ว่า: “เรามักใช้สำนวน นี่คือความงามแบบรัสเซียล้วนๆ นี่คือรูปกระต่ายถ่มน้ำลาย ซึ่งเป็นหน้าแบบรัสเซียทั่วไป เราสามารถมั่นใจได้ว่าไม่ใช่สิ่งที่น่าอัศจรรย์ แต่จริงอยู่ในนิพจน์ทั่วไปนี้ โหงวเฮ้งรัสเซีย. ในพวกเราแต่ละคนในขอบเขตของ "หมดสติ" ของเรามีแนวคิดที่ค่อนข้างแน่นอนของประเภทรัสเซีย "(A.P. Bogdanov" Anthropological Physiognomy ". M. , 1878)
หนึ่งร้อยปีต่อมาและตอนนี้นักมานุษยวิทยาสมัยใหม่ V. Deryabin โดยใช้วิธีการล่าสุดของการวิเคราะห์หลายมิติทางคณิตศาสตร์ของคุณสมบัติผสมมาถึงข้อสรุปเดียวกัน: “ข้อสรุปแรกและที่สำคัญที่สุดคือการตรวจสอบความสามัคคีที่สำคัญของรัสเซียทั่วรัสเซียและ ความเป็นไปไม่ได้ที่จะแยกแยะได้แม้กระทั่งประเภทภูมิภาคที่เกี่ยวข้อง แยกจากกันอย่างชัดเจน” (“Issues of Anthropology”, Issue 88, 1995) ความเป็นเอกภาพทางมานุษยวิทยารัสเซียนี้แสดงออกถึงความเป็นเอกภาพของลักษณะทางพันธุกรรมที่แสดงออกในรูปลักษณ์ของบุคคลในโครงสร้างของร่างกายของเขา?
ก่อนอื่น - สีผมและสีตารูปร่างของโครงสร้างของกะโหลกศีรษะ ตามคุณสมบัติเหล่านี้ เราชาวรัสเซียต่างจากชนชาติยุโรปและมองโกลอยด์ และเราไม่สามารถเทียบกับนิโกรและเซมิติได้เลย ความแตกต่างนั้นโดดเด่นเกินไปนักวิชาการ Alekseev พิสูจน์ความคล้ายคลึงกันในระดับสูงในโครงสร้างของกะโหลกศีรษะในบรรดาตัวแทนทั้งหมดของชาวรัสเซียสมัยใหม่ในขณะที่ระบุว่า "ประเภทโปรโต - สลาฟ" มีความเสถียรมากและมีรากฐานมาจากยุคหินใหม่และอาจเป็นหิน ตามการคำนวณของนักมานุษยวิทยา Deryabin ดวงตาสีอ่อน (สีเทา สีเทาสีน้ำเงิน สีฟ้า และสีน้ำเงิน) พบได้ในรัสเซีย 45 เปอร์เซ็นต์ ในยุโรปตะวันตกมีเพียง 35 เปอร์เซ็นต์เท่านั้นที่มีตาสว่าง ผมสีดำเข้มพบในรัสเซีย 5 เปอร์เซ็นต์ในประชากรของยุโรปต่างประเทศ - ใน 45 เปอร์เซ็นต์ ภูมิปัญญาดั้งเดิมเกี่ยวกับ "ความเย่อหยิ่ง" ของรัสเซียไม่ได้รับการยืนยันเช่นกัน ใน 75% ของชาวรัสเซีย พบว่ามีจมูกตรง
บทสรุปของนักมานุษยวิทยา:
“ชาวรัสเซียในองค์ประกอบทางเชื้อชาติของพวกเขาคือคอเคซอยด์ทั่วไป โดยครอบครองตำแหน่งศูนย์กลางในหมู่ประชาชนของยุโรปโดยลักษณะทางมานุษยวิทยาส่วนใหญ่และมีความโดดเด่นด้วยสีที่ค่อนข้างสว่างของดวงตาและผม นอกจากนี้ยังควรตระหนักถึงความสามัคคีที่สำคัญของประเภทเชื้อชาติของรัสเซียทั่วยุโรปรัสเซีย
“รัสเซียเป็นชาวยุโรป แต่เป็นชาวยุโรปที่มีลักษณะทางกายภาพเฉพาะสำหรับเขาเท่านั้น สัญญาณเหล่านี้ประกอบขึ้นเป็นสิ่งที่เราเรียกว่ากระต่ายทั่วไป
นักมานุษยวิทยาเการัสเซียอย่างจริงจังและ - ไม่มีตาตาร์นั่นคือมองโกลอยด์ในรัสเซีย หนึ่งในสัญญาณทั่วไปของมองโกลอยด์คือ epicanthus - รอยพับของมองโกเลียที่มุมด้านในของดวงตา ในมองโกลอยด์ทั่วไป พบรอยพับนี้ใน 95 เปอร์เซ็นต์ ในการศึกษาชาวรัสเซียแปดพันห้าพันคน พบรอยพับดังกล่าวใน 12 คนเท่านั้น และอยู่ในรูปของตัวอ่อน
ตัวอย่างอื่น. ชาวรัสเซียมีเลือดพิเศษอย่างแท้จริง - ความเด่นของกลุ่มที่ 1 และ 2 ซึ่งเห็นได้จากการปฏิบัติของสถานีถ่ายเลือดในระยะยาว ตัวอย่างเช่น ในชาวยิว กรุ๊ปเลือดที่โดดเด่นคืออันดับที่ 4 และปัจจัย Rh เชิงลบนั้นพบได้บ่อยกว่า ในการตรวจเลือดทางชีวเคมี ปรากฎว่ารัสเซีย เช่นเดียวกับคนยุโรปทั้งหมด มีลักษณะพิเศษของยีน RN-c ยีนนี้ไม่มีอยู่ใน Mongoloids (OV Borisova "Polymorphism ของกรดเม็ดเลือดแดง phosphatase ในกลุ่มประชากรต่างๆ สหภาพโซเวียต" "ประเด็นมานุษยวิทยาฉบับที่ 53, 2519)
ปรากฎว่าไม่ว่าคุณจะขูดรัสเซียด้วยวิธีใด คุณจะไม่พบตาตาร์ คุณจะไม่พบใครในตัวเขา สิ่งนี้ได้รับการยืนยันจากสารานุกรม "ประชาชนของรัสเซีย" ในบท "องค์ประกอบทางเชื้อชาติของประชากรรัสเซีย" ระบุไว้ว่า: "ตัวแทนของเผ่าพันธุ์คอเคซอยด์คิดเป็นมากกว่า 90 เปอร์เซ็นต์ของประชากรของประเทศและประมาณ 9 เปอร์เซ็นต์เป็น ตัวแทนของรูปแบบผสมระหว่างคอเคซอยด์และมองโกลอยด์ จำนวน Mongoloids บริสุทธิ์ไม่เกิน 1 ล้านคน (“ประชาชนของรัสเซีย”. M. , 1994)
เป็นเรื่องง่ายที่จะคำนวณว่าหากมีชาวรัสเซียร้อยละ 84 ในรัสเซีย แสดงว่าพวกเขาทั้งหมดเป็นคนประเภทยุโรปเท่านั้น ชาวไซบีเรีย, ภูมิภาคโวลก้า, คอเคซัส, เทือกเขาอูราลเป็นส่วนผสมของเชื้อชาติยุโรปและมองโกเลีย สิ่งนี้แสดงออกอย่างสวยงามโดยนักมานุษยวิทยา A.P. Bogdanov ในศตวรรษที่ 19 ศึกษาชนชาติของรัสเซียเขาเขียนโดยปฏิเสธจากตำนานปัจจุบันที่ห่างไกลและห่างไกลของเขาที่ชาวรัสเซียหลั่งเลือดจากต่างประเทศเข้าสู่ประชาชนของพวกเขาในยุคของการรุกรานและการล่าอาณานิคม:
“ชาวรัสเซียหลายคนอาจแต่งงานกับผู้หญิงพื้นเมืองและตั้งรกราก แต่อาณานิคมรัสเซียดั้งเดิมส่วนใหญ่ทั่วรัสเซียและไซบีเรียไม่เป็นเช่นนั้น เป็นการค้าขาย คนอุตสาหกรรม กระตือรือร้นที่จะจัดตัวเองตามแบบฉบับของตนเอง ตามอุดมคติของตนให้เกิดความอยู่ดีมีสุข และอุดมคติของคนรัสเซียนี้ไม่ได้เป็นเช่นนั้นเลยที่จะทำให้ชีวิตของเขาบิดเบี้ยวด้วย "ขยะ" บางอย่างได้ง่ายเนื่องจากตอนนี้คนรัสเซียให้เกียรติผู้ที่ไม่เชื่อบ่อยครั้ง เขาจะทำธุรกิจกับเขา จะรักใคร่และเป็นมิตรกับเขา จะเป็นเพื่อนกับเขาในทุกสิ่ง ยกเว้นการแต่งงาน เพื่อแนะนำองค์ประกอบแปลกปลอมเข้ามาในครอบครัวของเขา คนรัสเซียทั่วไปยังคงแข็งแกร่งในเรื่องนี้ และเมื่อพูดถึงครอบครัว จนถึงการหยั่งรากจากบ้านของพวกเขา ที่นี่เขามีชนชั้นสูง บ่อยครั้งที่ผู้ตั้งถิ่นฐานของชนเผ่าต่าง ๆ อาศัยอยู่ในละแวกนั้น แต่การแต่งงานระหว่างพวกเขานั้นหายาก
เป็นเวลาหลายพันปีที่รูปแบบทางกายภาพของรัสเซียยังคงมีเสถียรภาพและไม่เปลี่ยนแปลง และไม่เคยมีการผสมข้ามระหว่างชนเผ่าต่าง ๆ ที่อาศัยอยู่ในดินแดนของเราเป็นครั้งคราว ตำนานได้ถูกกำจัดไปแล้ว เราต้องเข้าใจว่าการเรียกร้องของเลือดไม่ใช่วลีที่ว่างเปล่า ว่าความคิดระดับชาติของเราเกี่ยวกับประเภทรัสเซียคือความเป็นจริงของสายพันธุ์รัสเซีย เราต้องเรียนรู้ที่จะเห็นสายพันธุ์นี้ ชื่นชม ชื่นชมในญาติชาวรัสเซียที่ใกล้ชิดและห่างไกลของเรา และบางทีรัสเซียอาจดึงดูดมนุษย์ต่างดาวโดยสิ้นเชิง แต่คนของเราสำหรับเราจะได้รับการฟื้นฟู - พ่อ แม่ พี่ชาย น้องสาว ลูกชายและลูกสาว ท้ายที่สุดแล้ว เราทุกคนล้วนมาจากรากเดียวกัน จากชนิดเดียว - ชนิดรัสเซีย
3) นักมานุษยวิทยาสามารถระบุลักษณะของคนรัสเซียทั่วไปได้ ในการทำเช่นนี้พวกเขาต้องแปลภาพถ่ายทั้งหมดจากคลังภาพถ่ายของพิพิธภัณฑ์มานุษยวิทยาในระดับเดียวด้วยภาพเต็มหน้าและโปรไฟล์ของตัวแทนทั่วไปของประชากรในภูมิภาครัสเซียของประเทศและรวมเข้าด้วยกันใน รูม่านตาซ้อนทับกัน ภาพบุคคลสุดท้ายกลายเป็นภาพเบลอ แต่พวกเขาให้ความคิดเกี่ยวกับการปรากฏตัวของคนรัสเซียอ้างอิง นี่เป็นการค้นพบครั้งแรกที่น่าตื่นเต้นอย่างแท้จริง อันที่จริง ความพยายามที่คล้ายคลึงกันของนักวิทยาศาสตร์ชาวฝรั่งเศสนำไปสู่ผลลัพธ์ที่พวกเขาต้องซ่อนตัวจากพลเมืองในประเทศของพวกเขา: หลังจากหลายพันชุดกับรูปถ่ายอ้างอิง Jacques และ Marianne ที่ได้รับแล้วใบหน้าวงรีสีเทาที่ไม่มีใบหน้าก็มอง ภาพดังกล่าวแม้ในหมู่ชาวฝรั่งเศสที่อยู่ห่างไกลจากมานุษยวิทยามากที่สุดก็อาจทำให้เกิดคำถามที่ไม่จำเป็น: มีประเทศในฝรั่งเศสหรือไม่?
น่าเสียดายที่นักมานุษยวิทยาไม่ได้ไปไกลกว่าการสร้างภาพเหมือนของตัวแทนทั่วไปของประชากรรัสเซียในภูมิภาคต่าง ๆ ของประเทศและไม่ได้ซ้อนทับกันเพื่อให้ได้รูปลักษณ์ของคนรัสเซียอย่างแท้จริง ในท้ายที่สุด พวกเขาถูกบังคับให้ยอมรับว่าภาพถ่ายดังกล่าวอาจทำให้พวกเขาเดือดร้อนในที่ทำงาน อย่างไรก็ตาม ภาพร่าง "ระดับภูมิภาค" ของชาวรัสเซียได้รับการตีพิมพ์ในสื่อทั่วไปในปี 2545 เท่านั้นและก่อนหน้านั้นพวกเขาได้รับการตีพิมพ์ในฉบับเล็ก ๆ เฉพาะในสิ่งพิมพ์ทางวิทยาศาสตร์สำหรับผู้เชี่ยวชาญเท่านั้น ตอนนี้คุณสามารถตัดสินด้วยตัวคุณเองว่ามีความคล้ายคลึงกับ Ivanushka และ Marya ในโรงภาพยนตร์ทั่วไปเพียงใด
น่าเสียดายที่รูปถ่ายเอกสารเก่าขาวดำส่วนใหญ่เป็นใบหน้าของคนรัสเซียไม่อนุญาตให้เราถ่ายทอดความสูง ร่างกาย สีผิว ผมและดวงตาของคนรัสเซีย อย่างไรก็ตาม นักมานุษยวิทยาได้สร้างภาพพจน์ของชายและหญิงรัสเซีย มีรูปร่างปานกลางและสูงปานกลาง มีผมสีน้ำตาลอ่อน ตาสีอ่อน - สีเทาหรือสีน้ำเงิน ในระหว่างการวิจัยยังได้ภาพวาจาของชาวยูเครนทั่วไปอีกด้วย ยูเครนอ้างอิงแตกต่างจากรัสเซียเฉพาะในสีผิวผมและดวงตาของเขา - เขาเป็นสีน้ำตาลเข้มที่มีลักษณะปกติและดวงตาสีน้ำตาล จมูกดูแคลนกลายเป็นสิ่งที่ไม่เคยมีมาก่อนของชาวสลาฟตะวันออก (พบเพียง 7% ของรัสเซียและยูเครน) คุณลักษณะนี้เป็นเรื่องปกติสำหรับชาวเยอรมัน (25%)
4) ในปี 2543 มูลนิธิรัสเซียเพื่อการวิจัยขั้นพื้นฐานได้จัดสรรเงินประมาณครึ่งล้านรูเบิลจากกองทุนงบประมาณของรัฐเพื่อศึกษากลุ่มยีนของคนรัสเซีย เป็นไปไม่ได้ที่จะใช้โปรแกรมอย่างจริงจังด้วยเงินทุนดังกล่าว แต่มันเป็นจุดสังเกตมากกว่าแค่การตัดสินใจทางการเงิน ซึ่งบ่งบอกถึงการเปลี่ยนแปลงในลำดับความสำคัญทางวิทยาศาสตร์ของประเทศ เป็นครั้งแรกในประวัติศาสตร์ของรัสเซียที่นักวิทยาศาสตร์จากห้องปฏิบัติการพันธุศาสตร์ประชากรมนุษย์ของศูนย์พันธุศาสตร์การแพทย์ของสถาบันวิทยาศาสตร์การแพทย์แห่งรัสเซียซึ่งได้รับทุนสนับสนุนจากมูลนิธิรัสเซียเพื่อการวิจัยขั้นพื้นฐานสามารถมีสมาธิในการศึกษายีนได้อย่างเต็มที่ กลุ่มคนรัสเซียและไม่ใช่คนตัวเล็กเป็นเวลาสามปี และเงินทุนที่จำกัดก็กระตุ้นความเฉลียวฉลาดของพวกเขาเท่านั้น พวกเขาเสริมการศึกษาอณูพันธุศาสตร์ด้วยการวิเคราะห์การกระจายความถี่ของนามสกุลรัสเซียในประเทศ วิธีนี้ราคาถูกมาก แต่เนื้อหาข้อมูลนั้นเกินความคาดหมายทั้งหมด: การเปรียบเทียบภูมิศาสตร์ของนามสกุลกับภูมิศาสตร์ของเครื่องหมาย DNA ทางพันธุกรรมแสดงให้เห็นความบังเอิญเกือบทั้งหมด
น่าเสียดายที่การตีความการวิเคราะห์ครอบครัวที่ปรากฏในสื่อหลังจากการตีพิมพ์ข้อมูลครั้งแรกในวารสารทางวิทยาศาสตร์เฉพาะทางอาจสร้างความประทับใจที่ผิดพลาดเกี่ยวกับเป้าหมายและผลลัพธ์ของงานใหญ่ของนักวิทยาศาสตร์ ผู้จัดการโครงการ Doctor of Sciences Elena Balanovskaya อธิบายว่าสิ่งสำคัญไม่ใช่ว่านามสกุล Smirnov กลายเป็นเรื่องธรรมดาในหมู่คนรัสเซียมากกว่า Ivanov แต่เป็นครั้งแรกที่มีการรวบรวมรายชื่อนามสกุลรัสเซียอย่างแท้จริงตามภูมิภาค ของประเทศ. อันดับแรก รายชื่อถูกรวบรวมสำหรับภูมิภาคที่มีเงื่อนไขห้าแห่ง ได้แก่ ภาคเหนือ ภาคกลาง ภาคกลาง-ตะวันตก ภาคกลาง-ตะวันออก และภาคใต้ โดยรวมแล้วมีการรวบรวมนามสกุลรัสเซียประมาณ 15,000 สกุลในทุกภูมิภาคซึ่งส่วนใหญ่พบได้ในภูมิภาคใดภูมิภาคหนึ่งเท่านั้นและไม่มีในภูมิภาคอื่น เมื่อรายการระดับภูมิภาคซ้อนทับกัน นักวิทยาศาสตร์ระบุชื่อสกุลทั้งหมด 257 รายการที่เรียกว่า "นามสกุลรัสเซียทั้งหมด" ที่น่าสนใจในขั้นตอนสุดท้ายของการศึกษาพวกเขาตัดสินใจที่จะเพิ่มชื่อของผู้อยู่อาศัยในดินแดนครัสโนดาร์ในรายการภาคใต้โดยคาดหวังว่าความเด่นของนามสกุลยูเครนของลูกหลานของคอสแซค Zaporizhzhya ขับไล่ที่นี่โดย Catherine II ลดรายชื่อรัสเซียทั้งหมดลงอย่างมาก แต่ข้อ จำกัด เพิ่มเติมนี้ลดรายชื่อนามสกุลรัสเซียทั้งหมดเพียง 7 หน่วย - เป็น 250 จากนั้นตามที่เห็นได้ชัดเจนและไม่ใช่สำหรับทุกคน ข้อสรุปที่น่าพอใจว่า Kuban ส่วนใหญ่อาศัยอยู่โดยคนรัสเซีย และชาวยูเครนไปไหนและอยู่ที่นั่นด้วย คำถามสำคัญคือ
เป็นเวลาสามปีที่ผู้เข้าร่วมโครงการ Russian Gene Pool ได้ใช้หลอดฉีดยาและหลอดทดลองเกือบทั่วทั้งอาณาเขตยุโรปของสหพันธรัฐรัสเซีย และทำตัวอย่างเลือดรัสเซียที่เป็นตัวแทนอย่างมาก
อย่างไรก็ตาม วิธีทางอ้อมราคาถูกสำหรับการศึกษาพันธุศาสตร์ของคนรัสเซีย (โดยใช้นามสกุลและ dermatoglyphics) เป็นเพียงส่วนเสริมสำหรับการศึกษาครั้งแรกในรัสเซียเกี่ยวกับกลุ่มยีนของสัญชาติที่มียศัยย์ ผลลัพธ์ทางพันธุกรรมระดับโมเลกุลหลักของเขามีอยู่ในเอกสาร Russian Gene Pool (Luch ed.) น่าเสียดาย เนื่องจากขาดเงินทุนของรัฐ นักวิทยาศาสตร์จึงต้องดำเนินการส่วนหนึ่งของการศึกษาร่วมกับเพื่อนร่วมงานชาวต่างชาติ ซึ่งกำหนดให้มีการเลื่อนการชำระหนี้หลายผลลัพธ์ จนกว่าจะมีการตีพิมพ์สิ่งพิมพ์ร่วมกันในสื่อทางวิทยาศาสตร์ ไม่มีอะไรขัดขวางเราจากการอธิบายข้อมูลเหล่านี้เป็นคำพูด ตามโครโมโซม Y ระยะห่างทางพันธุกรรมระหว่างชาวรัสเซียและฟินน์คือ 30 หน่วยตามแบบแผน และระยะห่างทางพันธุกรรมระหว่างคนรัสเซียกับคนที่เรียกว่า Finno-Ugric (Mari, Veps ฯลฯ ) ที่อาศัยอยู่ในอาณาเขตของสหพันธรัฐรัสเซียคือ 2-3 หน่วย พูดง่ายๆ ว่าพันธุกรรมเกือบจะเหมือนกัน ผลการวิเคราะห์ DNA ของไมโทคอนเดรียแสดงให้เห็นว่าชาวรัสเซียจากพวกตาตาร์อยู่ในระยะทางพันธุกรรมที่เท่ากันกับ 30 หน่วยทั่วไปที่แยกเราออกจากฟินน์ แต่ระหว่างชาวยูเครนจากลวิฟและตาตาร์นั้น ระยะห่างทางพันธุกรรมเพียง 10 หน่วย. และในขณะเดียวกัน ชาวยูเครนจากฝั่งซ้ายของยูเครนก็มีพันธุกรรมที่ใกล้เคียงกับชาวรัสเซียอย่าง Komi-Zyryans, Mordvins และ Mari
ภาษามียีนหรือไม่? เหตุใดยีนพูลจึงต้องการชื่อ - แผนที่ระยะทางพูดว่าอย่างไร - ไม่ใช่การ์ดใบเดียว แต่เป็นแฟนทั้งหมด!§หนึ่ง. ระยะทางจากตระกูลภาษาสามตระกูล:จากอินโด-ยุโรป: ความแตกต่างเพิ่มขึ้นทางทิศตะวันออก - แต่ประชากรส่วนใหญ่มีความใกล้ชิดทางพันธุกรรม - จาก URAL: ระยะทางเพิ่มขึ้นจากตะวันออกไปตะวันตก - แต่มีประชากรจำนวนมากอยู่ใกล้ - ชั้นล่าง Finno-Ugric ในหมู่ Slavs และ Turks; - จากอัลไต: ใกล้ชิดกับตนเองเท่านั้น - ไม่มีอิทธิพลต่อเพื่อนบ้านในยุโรป
§2. ระยะทางจากรัสเซีย, เบลารุส, ยูเครน:เครื่องหมายคลาสสิก - รัสเซียเหนืออยู่ไกลจากรัสเซียโดยเฉลี่ยมากกว่ายูเครนมอร์โดเวียนและชูวัช - เครื่องหมายดีเอ็นเอออโตโซม - ภาพเก่า - เกือบทุกคนอยู่ใกล้กับรัสเซีย - ยกเว้นคอเคซัสและอูราล - โครโมโซม Y - ภาพเดียวกันที่มีความเปรียบต่างมากกว่า - ระยะทางจากเบลารุส - คล้ายกันสำหรับชาวสลาฟเท่านั้น - ภาพเดียวกันสำหรับชาวยูเครน - ดังนั้นประชากรยุโรปตะวันออกจึงอยู่ใกล้กับรัสเซียและไม่ใช่ชาวสลาฟโดยทั่วไป!
ภาษามียีนหรือไม่?
เราต้องการตอบทันทีว่าผู้เขียนเช่นผู้อ่านรู้ว่าภาษาไม่มียีน สิ่งนี้สามารถเข้าใจได้แม้ในชีวิตประจำวัน - มีชาวรัสเซียกี่คนที่กระจัดกระจายไปทั่วโลกโดยคลื่นของการอพยพครั้งแรกครั้งที่สองและอื่น ๆ พูดหลายภาษา! และมียีนที่สืบทอดมาจากบรรพบุรุษเดียวกัน
แล้วทำไมเราถึงพูดถึงยีนของตระกูลภาษาสลาฟหรือภาษาเยอรมัน? มันเป็นวิทยาศาสตร์? ค่อนข้าง. ท้ายที่สุดเรามีส่วนร่วมในพันธุศาสตร์ของประชากรและเรากำลังพูดถึงเฉพาะประชากรของผู้ที่พูดภาษาของสาขาภาษาสลาฟหรือภาษาเยอรมันเท่านั้น และไม่มีอะไรอื่นอยู่เบื้องหลัง "ชื่อภาษาศาสตร์"
เราได้พูดไปแล้วหลายครั้งว่าประชากรมีหลายชั้นและอาจมีระดับที่แตกต่างกันมาก - ตั้งแต่ประชากรเบื้องต้น (หมู่บ้านใกล้เคียงหลายแห่ง) ไปจนถึงประชากรของมนุษยชาติทั้งหมด ทั้งหมดนี้เป็นประชากร และพวกมันซ้อนกันเหมือนตุ๊กตาทำรัง: ประชากรที่มีตำแหน่งต่ำกว่าจำนวนมากพอดีกับประชากรในระดับที่สูงกว่าถัดไป และอื่นๆ เรากำหนดหนึ่งในประชากรขั้นกลางเหล่านี้โดยคร่าวๆ ตามเชื้อชาติ นี่เป็นเหตุผลเดียวที่เราสามารถพูดถึงกลุ่มยีนของรัสเซียได้ นั่นคือ เกี่ยวกับประชากรที่ทำเครื่องหมายว่าเป็นของคนรัสเซีย ยิ่งกว่านั้นสิ่งของนี้ถูกกำหนดโดยผู้คนเองและไม่ว่าในกรณีใดโดยพันธุกรรม! และหลังจากที่ผู้คนระบุว่าตนเองเป็นชาวรัสเซียหรือชาวนอร์เวย์ (หรือรายงานว่าปู่ย่าตายายของพวกเขาคิดเกี่ยวกับเรื่องนี้) นักพันธุศาสตร์ก็เริ่มดูไม่ลำเอียง: ทำและประชากรของรัสเซียและนอร์เวย์แตกต่างกันมากน้อยเพียงใด? เราเรียกประชากรเหล่านี้อย่างมีเงื่อนไขว่า "รัสเซีย" หรือ "นอร์เวย์" ในขณะที่ตระหนักดีว่ากลุ่มยีนและประชากรเป็นหน่วยทางชีววิทยาที่เราตั้งชื่อให้ "มนุษยธรรม"
แต่เราเน้นว่าการที่เราตั้งชื่อกลุ่มยีนว่า "รัสเซีย" หรือ "นอร์เวย์" ไม่ได้หมายความว่า "ยีนรัสเซีย" หรือ "ยีนนอร์เวย์" ปรากฏขึ้นในที่เกิดเหตุทันที! ไม่มียีน "รัสเซีย" หรือ "ยูเครน" เช่นเดียวกับที่ไม่มียีนสลาฟหรือโรมานซ์ ไม่สิ เพราะยีนนั้นแก่กว่าคนมากและกระจัดกระจายไปทั่วโลก อย่างไรก็ตาม เราพูดถึงประเด็นเหล่านี้ในช่วงท้ายของหนังสือ (บทที่ 10) และตอนนี้เป็นสิ่งสำคัญสำหรับเราเท่านั้นที่จะตอบคำถาม - หากไม่มียีนรัสเซียหรือสลาฟทำไมเราถึงเรียกยีนพูลด้วยชื่อดังกล่าว
ทำไมยีนพูลชื่อ?
เพียงเพราะว่าประชากร (และกลุ่มยีนของพวกมัน) จะต้องได้รับชื่อที่มีความหมาย แน่นอนคุณสามารถออกจากสระยีนที่ไม่มีชื่อและทำซ้ำตลอดเวลา "ประชากรหลักในชนบทของภูมิภาคของที่ราบยุโรปตะวันออกและภูมิภาคทางเหนืออื่น ๆ มีความสัมพันธ์คร่าวๆกับพรมแดนของรัฐรัสเซียก่อน Ivan the Terrible ." แต่แม้จากวลีดังกล่าว ก็ยังไม่ชัดเจนว่าใครที่เรายังคงวิเคราะห์อยู่ (เช่น ไม่ว่าเราจะรวม Karelians, Izhora, Tatars หรือ Mordovians) และถ้าเราพูด (ตามที่อธิบายไว้ในรายละเอียดในตอนต้นของหนังสือ) ว่าภายใต้แหล่งพันธุกรรมของรัสเซีย เราจะหมายถึงชาวรัสเซียในชนบทที่เป็นชนพื้นเมืองในพื้นที่ "ดั้งเดิม" (ประวัติศาสตร์) แล้วใช้คำว่า "กลุ่มยีนรัสเซีย" ตลอด หนังสือจึงเป็นเรื่องง่ายสำหรับผู้อ่านจะเข้าใจสิ่งที่ผู้เขียนกำลังพูดถึง ดังนั้นเราจึงตั้งชื่อตามเงื่อนไขของกลุ่มยีน - เพื่อความสะดวกในการทำความเข้าใจ
อย่างไรก็ตาม เพื่อให้ชื่อแก่ตุ๊กตาทำรังที่มียศสูงกว่า ต้องใช้การจำแนกประเภทของประชากรบางประเภท ตัวอย่างเช่น ในบทที่ 2 เราได้ทดสอบว่าการจำแนกทางเชื้อชาติและภาษาศาสตร์มีประสิทธิภาพทางพันธุกรรมอย่างไร และในบรรดาชนชาติไซบีเรีย พวกเขาได้ทดสอบประสิทธิภาพทางพันธุกรรมของการจำแนกชนชาติตามประเภทของเครื่องประดับและประเภทของรำมะนาด และปรากฎว่าเครื่องประดับเผยให้เห็นประชากรได้ไม่ดี แต่แทมบูรีนหมอผีไม่ได้มีประสิทธิภาพในการแยกแยะประชากรน้อยกว่าภาษา แต่อย่างไรก็ตาม การจำแนกประเภททางภาษาศาสตร์ได้รับการพัฒนาอย่างละเอียดถี่ถ้วน นั่นคือเหตุผลที่ชื่อประชากรมักใช้ชื่อภาษาต่างๆ นี่คือวิธีที่เป็นอยู่ในปัจจุบันในวิทยาศาสตร์ชีวภาพ ตัวอย่างเช่นเมื่อเราพูดถึงชั้น "Finno-Ugric" ของแหล่งรวมยีน ทั้งนักมานุษยวิทยาและนักโบราณคดีก็เข้าใจเรา พวกเขาเข้าใจว่าเรากำลังพูดถึงประชากรจำนวนหนึ่ง ซึ่งค่อนข้างขยายเวลาและพื้นที่ออกไป และไม่สำคัญว่าตอนนี้ Chuvash ได้เปลี่ยนภาษาเดิมเป็นภาษาเตอร์กแล้ว และไม่สำคัญว่าเราไม่รู้ว่าภาษาที่ประชากรโบราณพูดภาษาใดหากพวกเขาไม่ทิ้งหลักฐานเป็นลายลักษณ์อักษรไว้ ข้อมูลจำนวนมหาศาลจากศาสตร์ต่างๆ (เช่น ชื่อแม่น้ำหรือทะเลสาบ) บ่งบอกว่ามีชุมชนคนอยู่ ซึ่งตอนนี้เราตั้งชื่อตามเงื่อนไขของโลก "Finno-Ugric"
ดังนั้นในเรื่องนี้และในหัวข้อถัดไป การเปรียบเทียบระยะทางทางพันธุกรรมจากประชากรที่มีชื่อ "ภาษาศาสตร์" เราไม่เปลี่ยนวิทยาศาสตร์ แต่ทำตามกฎที่เข้มงวด เราใช้การจำแนกภาษาของชนชาติ จากนั้น ตามนั้น เราให้ชื่อ "ภาษาศาสตร์" แบบมีเงื่อนไขแก่กลุ่มประชากรแต่ละกลุ่ม และสุดท้าย เราคำนวณความถี่ยีนเฉลี่ยสำหรับประชากรจากกลุ่มนี้ที่อาศัยอยู่ในอาณาเขตที่วิเคราะห์ จากนั้นเราจะดูว่าประชากรแต่ละกลุ่มในยุโรปตะวันออกแตกต่างจากความถี่เฉลี่ยเหล่านี้ของอาร์เรย์ประชากร "อินโด-ยูโรเปียน" หรือ "อัลไต" อย่างไร ในเวลาเดียวกัน ผู้เขียนและผู้อ่านต่างทราบดีว่าภาษาอัลไตอิกในยุโรปนั้นพูดโดยผู้คนที่มีรูปลักษณ์ที่แตกต่างกันโดยสิ้นเชิง ตั้งแต่ Gagauz ไปจนถึง Kalmyks แต่เราไม่มีสิทธิ์บนพื้นฐานนี้ที่จะกีดกันใครก็ตามจากกลุ่มที่ภาษาศาสตร์ได้ระบุ - เราระบุอย่างตรงไปตรงมาว่าคนใดบ้างที่รวมอยู่ในประชากรด้วยชื่อ "ภาษาศาสตร์" ที่กำหนด
บัตร DISTANCE พูดถึงอะไร?
แผนที่ระยะทางทางพันธุกรรมมีความสำคัญมากกว่าแผนที่องค์ประกอบหลัก เครื่องมือหลักสองอย่างนี้ของภูมิศาสตร์ยีน ใช้ร่วมกัน ให้คำอธิบายเสริมของพูลยีน แผนที่องค์ประกอบหลักช่วยให้เราสามารถเสนอสมมติฐานเกี่ยวกับปัจจัยที่ก่อให้เกิดรูปแบบที่สังเกตได้ และแผนที่ระยะทางทางพันธุกรรมช่วยให้เราสามารถทดสอบสมมติฐานเหล่านี้ได้
แต่ละแผนที่ของระยะทางทางพันธุกรรมที่นำเสนอในส่วนนี้เป็นค่าเฉลี่ยสำหรับตำแหน่งที่ศึกษาทั้งหมด (ตาราง 8.1.1) มันแสดงให้เห็นอย่างชัดเจนว่าพันธุกรรมแต่ละกลุ่มปิดกันอย่างไรในกลุ่มประชากรหนึ่งกลุ่มที่ผู้วิจัยระบุ กลุ่มประชากรดังกล่าวเรียกว่า "การอ้างอิง"
กลุ่มยีนสามารถถามคำถามได้: ประชากรกลุ่มใดที่มีความใกล้ชิดทางพันธุกรรมกับกลุ่มประชากรที่เราสนใจ ซึ่งค่อนข้างถูกลบออก? และสิ่งใดที่แตกต่างจากกลุ่มอ้างอิงโดยพื้นฐานยิ่งไปกว่านั้นในชุดความถี่ยีนทั้งหมด? และแผนที่ของระยะทางทางพันธุกรรมจะให้คำตอบว่าแต่ละจุดบนแผนที่นั้นอยู่ใกล้หรือไกลจากกลุ่มอ้างอิงมากเพียงใด เราจะเห็นมันด้วยตาเราเอง
แผนที่ระยะทางแตกต่างจากการใช้ระยะทางทางพันธุกรรมตามปกติในแผนที่เดียวเท่านั้น แต่คุณลักษณะที่สำคัญที่สุด: เมื่อทำแผนที่ การวิเคราะห์จะรวมถึงพื้นที่ของประชากร เช่น ภูมิศาสตร์ ด้านพื้นที่
แผนที่ระยะทางทางพันธุกรรมมักเปิดเผยความสัมพันธ์ระหว่างระยะทางทางพันธุกรรมกับระยะทางทางภูมิศาสตร์ แผนที่แสดงให้เห็นว่า เมื่อเราย้ายออกจากประชากรอ้างอิง (ระบุโดยผู้วิจัย) ประชากรของพื้นที่ใกล้เคียงและดินแดนห่างไกลมากขึ้นจะมีความแตกต่างทางพันธุกรรมจากประชากรอ้างอิงมากขึ้นเรื่อยๆ อย่างไร อย่างไรก็ตาม ระยะทางทางพันธุกรรมที่เพิ่มขึ้นนี้ไม่ได้ขึ้นกับความห่างไกลทางภูมิศาสตร์เท่านั้น มิฉะนั้น แผนที่ของระยะทางทางพันธุกรรมใดๆ จะประกอบด้วยวงกลมที่มีจุดศูนย์กลาง เช่น วงกลมที่แยกจากหินที่ขว้างไปบนน้ำ
ในความเป็นจริง ระยะทางในบางทิศทางอาจเพิ่มขึ้นอย่างรวดเร็ว ซึ่งบ่งชี้ถึงอุปสรรคต่อการไหลของยีน ในอีกทางหนึ่ง ระยะทางแทบจะไม่เพิ่มขึ้นเลย ซึ่งแสดงถึงความใกล้ชิดทางพันธุกรรมของกลุ่มที่อยู่ติดกันเหล่านี้ ในบางกรณี ความราบรื่นของไอโซลีนอาจถูกรบกวน และในหมู่กลุ่มที่ใกล้ชิดทางพันธุกรรม มีการตรวจพบประชากรที่อยู่ห่างไกลจากพันธุกรรม ซึ่งอาจบ่งบอกถึงการอพยพไปยังดินแดนนี้ ตัวอย่างเช่น ดังนั้น การวางแผนระยะทางทางพันธุกรรมบนแผนที่สามารถให้ข้อมูลที่มีค่าเกี่ยวกับความสัมพันธ์ของกลุ่มที่ศึกษากับประชากรอื่นๆ ในภูมิภาค การมีอยู่ของกระแสพันธุกรรม อุปสรรคทางพันธุกรรม และกลุ่มที่เกี่ยวข้อง นอกจากนี้เรายังได้รับข้อมูลเกี่ยวกับกลุ่มอ้างอิง (เช่น รัสเซียหรือเบลารุส): เกี่ยวกับความหลากหลายทางพันธุกรรมภายใน เกี่ยวกับการเบี่ยงเบนจากค่าเฉลี่ยภายในช่วงของมันเอง
ไม่ใช่บัตรเดียว แฟนทั้งหมด!
การทำแผนที่ระยะทางทางพันธุกรรมทำให้เห็นคุณลักษณะหลายอย่างของแหล่งรวมยีน โดยเฉพาะอย่างยิ่งหากเราพิจารณาไม่ใช่แผนที่ระยะทางเดียว (จากคนคนเดียว) แต่เป็นแผนที่ชุดหนึ่ง - จากชนชาติต่างๆ จากกลุ่มประชากรหลัก แผนที่ใหม่แต่ละแผนที่จะบอกตำแหน่งทางพันธุกรรมของคนกลุ่มใหม่หรือกลุ่มประชาชนในกลุ่มยีนทั่วไปของภูมิภาค การเปรียบเทียบผู้ชื่นชอบแผนที่ทั้งหมดจะแสดงให้เห็นว่าแต่ละกลุ่มมีส่วนร่วมกับกลุ่มยีนยุโรปตะวันออกได้ดีเพียงใด และโซนของการผสมอยู่ตรงไหน
เราจะไม่พิจารณาแผนที่ของระยะทางทางพันธุกรรมจากผู้คนในยุโรปตะวันออกที่นี่ มิฉะนั้น เราจะไปไกลเกินกว่าขอบเขตของหนังสือเกี่ยวกับกลุ่มยีนรัสเซีย แผนที่ระยะทางจากกลุ่มคนที่เกี่ยวข้องกันมีข้อมูลมากขึ้น พวกเขาเปิดเผยรูปแบบที่ไม่เกี่ยวกับชาติพันธุ์ของชนชาติใด ๆ แต่เกี่ยวกับเหตุการณ์ทั่วไปในการก่อตัวของประชากรของยุโรปตะวันออก ดังที่เราได้พูดคุยกันในบทที่ 2 ลำดับวงศ์ตระกูลจะขึ้นอยู่กับหลักการของ "การปรับขนาด": เมื่อขนาดของกลุ่มที่ศึกษาเพิ่มขึ้น ร่องรอยของเหตุการณ์โบราณและขนาดใหญ่มากขึ้นเรื่อยๆ ก็ถูกเปิดเผย
ดังนั้นจึงให้ความสนใจกับแผนที่ระยะทางจากกลุ่มคน ในหัวข้อที่ 1 แผนที่จะถูกสร้างขึ้นจากความถี่เฉลี่ยของยีนในกลุ่มชนชาติตระกูลภาษาอินโด-ยูโรเปียน อูราลิก และอัลไต ที่อาศัยอยู่ในยุโรปตะวันออก จากนั้น (§2) เราพิจารณาแผนที่ระยะทางจากคนรัสเซีย โดยแสดงตำแหน่งของพวกเขาในกลุ่มยีนทั่วไปของยุโรปตะวันออก และโดยสรุป มาดูแผนที่ของชาวยุโรปตะวันออกอีกสองคนคือ เบลารุสและยูเครน ซึ่งในอดีตเคยใกล้ชิดกับประชากรรัสเซียและอาจมีกลุ่มยีนที่คล้ายคลึงกัน
บัตรทั้งหมดจะถูกอ่านในลักษณะเดียวกัน ยิ่งจุดนี้ของแผนที่ถูกลบออกจากกลุ่มประชากรอ้างอิงมาก ระยะทางยิ่งไกล สีของจุดนี้ก็จะยิ่งเข้มขึ้น ดังนั้นบริเวณที่เบาที่สุดจึงเป็นพื้นที่ที่มีระยะทางน้อยที่สุด เหล่านี้เป็นประชากรที่ใกล้เคียงกับการอ้างอิงมากที่สุด ที่มืดที่สุดคือบริเวณที่มีระยะทางไกลที่สุด เหล่านี้เป็นประชากรที่ไม่คล้ายคลึงกับข้อมูลอ้างอิงทางพันธุกรรม แน่นอน. ทันทีที่เราใช้ประชากรอ้างอิงที่แตกต่างกัน จุดเดียวกันบนแผนที่จะรายงานว่ามีระยะทางที่แตกต่างกันไปจากเกณฑ์มาตรฐานใหม่ เพื่อความสะดวกในการอ่าน แผนที่ระยะทางทั้งหมดสร้างขึ้นในระดับเดียว ดังนั้นคุณจึงสามารถเปรียบเทียบได้อย่างปลอดภัย ไม่เพียงแต่ส่วนต่างๆ ของแผนที่เดียว แต่ยังรวมถึงแผนที่ที่แตกต่างกันในแง่ของความเข้มของสี
§หนึ่ง. ระยะทางจากตระกูลภาษาสามภาษา
พิจารณาแผนที่ระยะทางทางพันธุกรรมของประชากรทั้งหมดในยุโรปตะวันออกจากชนชาติในตระกูลภาษาอินโด-ยูโรเปียน อูราลิก และอัลไตที่อาศัยอยู่ เพื่อความกระชับ เราขอนำเสนอแผนที่ของ "ผู้เห็นเหตุการณ์" คนหนึ่ง - เครื่องหมายดีเอ็นเอ autosomal เนื่องจากแผนที่ของระยะทางทางพันธุกรรมสำหรับเครื่องหมายแบบคลาสสิก ดังที่เราเห็นในย่อหน้าถัดไป ค่อนข้างคล้ายกัน
จากคนในครอบครัวภาษาอินโด-ยุโรป (DNA MARKERS)
แผนที่ระยะทางทางพันธุกรรมจากตระกูลภาษาอินโด - ยูโรเปียนแสดงในรูปที่ 8.3.1.
แผนที่ถูกสร้างขึ้นเช่นนี้ ประการแรก ความถี่เฉลี่ยของเครื่องหมายดีเอ็นเอถูกคำนวณสำหรับตัวแทนของตระกูลอินโด-ยูโรเปียนในยุโรปตะวันออก: ประชากรของรัสเซีย, ยูเครน, เบลารุส, มอลโดวา จากนั้นจึงหาความถี่ยีน "อินโด - ยูโรเปียน" โดยเฉลี่ย นอกจากนี้ ระยะทางทางพันธุกรรมจากความถี่ "อินโด-ยูโรเปียน" เฉลี่ยเหล่านี้ไปยังความถี่ที่จุดแต่ละจุดของแผนที่จะถูกคำนวณ และค่าระยะทางที่ได้รับจะอยู่ในโหนดเดียวกันของแผนที่
ดังนั้น ตัวอย่างเช่น หากในเบลารุสส่วนใหญ่ ในภูมิภาค Kyiv และ Lvov ค่าของระยะทางทางพันธุกรรมอยู่ในช่วง 0.01 ถึง 0.02 (รูปที่ 8.3.1.) ซึ่งหมายความว่าสิ่งเหล่านี้คือ ความแตกต่าง (โดยเฉลี่ยสำหรับยีนทั้งหมด) ของประชากรเหล่านี้จากความถี่กลางของชาวตระกูลอินโด-ยูโรเปียน ในทางตรงกันข้ามความแตกต่างระหว่าง Kalmyks, Komi และ Bashkirs นั้นยิ่งใหญ่กว่ามาก - ค่าของระยะทางทางพันธุกรรมในดินแดนของการตั้งถิ่นฐานของพวกเขามากกว่า 0.05 และ 0.06 แผนที่อื่น ๆ ของระยะทางทางพันธุกรรมอ่านในทำนองเดียวกัน
แผนที่แสดงให้เห็นว่าตามที่คาดไว้ ประชากรของชาวรัสเซียในรัสเซียกลาง ยูเครน เบลารุส มอลโดวา (นั่นคือประชากรอินโด-ยูโรเปียนเอง) ใกล้เคียงกับความถี่เฉลี่ยของชาวอินโด-ยูโรเปียนในยุโรปตะวันออก อย่างไรก็ตาม ไม่ใช่ทุกสิ่งที่สามารถคาดเดาได้ - ประชากรรัสเซียตอนเหนือ (แม้ว่าพวกเขาจะเป็นชาวอินโด-ยูโรเปียน) นั้นแตกต่างอย่างเห็นได้ชัดจาก "ชาวอินโด-ยูโรเปียนตอนกลาง" ในระดับเดียวกับชนชาติที่ไม่ใช่ชาวอินโด-ยูโรเปียนของแม่น้ำโวลก้าตอนกลาง (Mari, Mordovians, Chuvash) และคอเคซัสตะวันตก ในที่สุดประชากรของเทือกเขาอูราล (โดยเฉพาะชาวโคมิ) รวมถึงชนชาติบริภาษ (บัชคีร์, คาลมิกส์) กลับกลายเป็นสิ่งที่แตกต่างกันมากที่สุด
ให้ความสนใจกับประชากรชาวรัสเซีย พวกเขาเป็นตัวแทนของตระกูลภาษาอินโด-ยูโรเปียนในยุโรปตะวันออก และความถี่ของพวกมันถูกใช้ในการคำนวณความถี่ "อินโด-ยูโรเปียน" โดยเฉลี่ย และถึงกระนั้น เราเห็นความแตกต่างที่ชัดเจนระหว่างประชากรรัสเซียในแง่ของระดับความใกล้ชิดกับประชากรอ้างอิงของพวกเขาเอง สิ่งนี้บ่งชี้อีกครั้งว่าระดับของความหลากหลายในกลุ่มยีนของคนรัสเซียนั้นยิ่งใหญ่มากจนแสดงออกได้อย่างชัดเจนแม้ในระดับยุโรปตะวันออก
โดยทั่วไปมีการเปิดเผยรูปแบบทางภูมิศาสตร์ที่ชัดเจน: เมื่อเคลื่อนตัวไปทางทิศตะวันออก ค่าของระยะทางจะค่อยๆ เพิ่มขึ้น ประชากรมีความแตกต่างกันมากขึ้นเรื่อยๆ จากลักษณะทั่วไปของชาวอินโด-ยูโรเปียน และประชาชนในเขตชานเมืองทางตะวันออกของยุโรป กลับกลายเป็นว่ามีความแตกต่างทางพันธุกรรมมากที่สุดสำหรับพวกเขา อย่างไรก็ตาม โดยทั่วไปแล้ว คนส่วนใหญ่ในยุโรปตะวันออก (รวมถึงประชากรอูราลและคอเคเซียน) อยู่ใกล้กับชนชาติอินโด-ยูโรเปียน: ค่าเฉลี่ยของระยะทางทางพันธุกรรมบนแผนที่มีค่าน้อย d=0.028
จากคนในครอบครัวภาษาอูราล (เครื่องหมายดีเอ็นเอ)
แผนที่ระยะทางทางพันธุกรรมถัดไปสร้างขึ้นจากความถี่เฉลี่ยของยีนของตระกูลภาษาอูราลิกและแสดงภาพที่แตกต่าง (รูปที่ 8.3.2)
ในตระกูลอูราล มีเพียงชนชาติที่พูดภาษาฟินแลนด์ตะวันออก (Komi, Udmurts, Mari, Mordvins) เท่านั้นที่ได้รับการศึกษาสำหรับเครื่องหมายดีเอ็นเอ ระยะทางขั้นต่ำพบได้ในอาณาเขตของการตั้งถิ่นฐานของชนชาติเหล่านี้โดยเฉพาะในเทือกเขาอูราล ในทางตรงกันข้ามประชากรทางตะวันตกของที่ราบรัสเซียและ Ciscaucasia ถูกกำจัดทางพันธุกรรมออกจากความถี่อูราลโดยเฉลี่ย ภูมิภาคมัธยฐานของยุโรปตะวันออกซึ่งอยู่ติดกับเทือกเขาอูราลนั้นอยู่ใกล้กับชนชาติอูราลและในเชิงพันธุกรรม
ดังนั้น, ค่าระยะทางที่น้อยที่สุดจะถูกแปลเป็นภาษาอูราลและไปทางตะวันตกค่อยๆเพิ่มขึ้นอาจเป็นไปได้ว่าดินแดนที่ถูกครอบครองโดยค่ากลางนั้นสะท้อนถึงช่วงของชนเผ่า Finno-Ugric โบราณที่หลอมรวมโดย Slavs [Alekseeva, 1965] อยากรู้ว่าช่วงของชนชาติที่พูดภาษาเตอร์กของเทือกเขาอูราลนั้นใกล้เคียงกับลักษณะของตระกูลอูราลซึ่งอธิบายได้จากส่วนแบ่งที่สำคัญของสารตั้งต้นอูราลในกลุ่มยีนของชูวัชตาตาร์และบางกลุ่ม Bashkirs (โรกินสกี้, เลวิน, 1978)
ค่าเฉลี่ยของระยะทางบนแผนที่ แม้ว่าจะสูงกว่าจาก "อินโด-ยูโรเปียน" แต่ก็ไม่ใหญ่นัก (d=0.039) สิ่งนี้ยืนยันการเป็นตัวแทนที่สำคัญของกลุ่มยีนที่พูดภาษาอูราลในกลุ่มยีนยุโรปตะวันออกทั่วไป ซึ่งส่วนใหญ่ประกอบด้วยสารตั้งต้นของอูราล
จากคนในครอบครัวภาษาอัลไต (DNA MARKERS)
แผนที่ต่อไปนี้ (รูปที่ 8.3.3) แสดงความแตกต่างระหว่างประชากรยุโรปตะวันออกแต่ละกลุ่มและกลุ่มชนในตระกูลภาษาอัลไตอิก ครอบครัวนี้ในยุโรปตะวันออกส่วนใหญ่เป็นชนชาติที่พูดภาษาเตอร์ก - มีเพียง Kalmyks เท่านั้นที่พูดภาษาที่เป็นของกลุ่มมองโกเลียของตระกูลนี้
แผนที่ระยะทางทางพันธุกรรมสองแผนที่ก่อนหน้านี้ (จากตระกูลอินโด - ยูโรเปียนและจากตระกูลอูราล) นั้นโดดเด่นด้วยระยะทางเฉลี่ยเล็กน้อย บนแผนที่ (รูปที่ 8.3.1., 8.3.2.) สิ่งนี้สังเกตเห็นได้ชัดเจนจากความเด่นของสีอ่อน ในทางตรงกันข้าม แผนที่ระยะทางจากตระกูลอัลไต (รูปที่ 8.3.3) ถูกครอบงำด้วยสีเข้ม ซึ่งสอดคล้องกับระยะห่างทางพันธุกรรมที่สำคัญของประชากรยุโรปตะวันออกส่วนใหญ่จากกลุ่มยีนของตระกูลภาษาศาสตร์อัลตาอิก เฉพาะพื้นที่ของคนในตระกูลภาษาอัลตาอิกเท่านั้นที่ใกล้เคียงกับค่าเฉลี่ยโดยธรรมชาติ และทันทีที่อยู่นอกเขตการตั้งถิ่นฐาน ประชากรยุโรปตะวันออกที่เหลือนั้นแตกต่างอย่างมากจากลักษณะทางพันธุกรรมของชนชาติที่พูดภาษาอัลไต
สิ่งนี้ยังสะท้อนให้เห็นในคุณค่าของระยะทางทางพันธุกรรมที่มากกว่าแผนที่ก่อนหน้านี้ โดยเฉลี่ยแล้ว พวกมันมีจำนวน d = 0.064 บนแผนที่ ซึ่งสูงกว่าค่าของชาวอินโด-ยูโรเปียนที่ใกล้เคียงกันเกือบสามเท่า
ดังนั้นอิทธิพลของผู้คนในตระกูลอัลไตต่อกลุ่มยีนยุโรปตะวันออกจึงถูก จำกัด โดยโซนของการตั้งถิ่นฐานเท่านั้นและตามข้อมูลที่อยู่ระหว่างการพิจารณานั้นแทบจะไม่ได้ติดตามแม้แต่ในดินแดนที่อยู่ติดกัน ข้อเท็จจริงนี้สามารถอธิบายได้ด้วยการปรากฏตัวที่ค่อนข้างช้าในยุโรปตะวันออกของหลายเผ่าที่พูดภาษาของตระกูลอัลไต [ผู้คนและศาสนาของโลก, 1999] ในขณะที่ทั้งตระกูลอินโด - ยูโรเปียนและอูราลิกเป็นภาษา ของประชากรโบราณของยุโรปตะวันออก [Cheboksarov, Cheboksarova, 1971; บุนัค, 1980].
§2. ระยะทางจากรัสเซีย, เบลารุส, ยูเครน
ดังนั้นเราจึงได้เรียนรู้ "องค์ประกอบ" หลักของกลุ่มยีนยุโรปตะวันออก - กลุ่มย่อยของยีนหลักที่แสดงอยู่ในนั้น ในสิ่งที่ "หุ้น" พวกเขา "ผสม" และการแบ่งปันเหล่านี้แตกต่างกันอย่างไรในส่วนต่างๆ ของยุโรปตะวันออก ตอนนี้ เราสามารถกลับไปที่หัวข้อหลักของหนังสือของเรา และพิจารณาว่าประชากรยุโรปตะวันออกทั้งหมดที่สัมพันธ์กับรัสเซียเป็นอย่างไร เนื่องจากหัวข้อนี้เป็นผู้นำ เราจะให้ระยะห่างทางพันธุกรรมจากประชากรรัสเซียสำหรับเครื่องหมายสามประเภท - เครื่องหมายคลาสสิก เครื่องหมายดีเอ็นเอ autosomal และเครื่องหมายโครโมโซม Y และเพื่อไม่ให้สับสนระหว่างคุณลักษณะ "รัสเซียล้วน" กับ "สลาฟ" เราจะพิจารณาแผนที่ระยะทางจากชนชาติสลาฟตะวันออกที่ใกล้ชิดในอดีต - เบลารุสและยูเครน
จากประชากรรัสเซีย (เครื่องหมายคลาสสิก)
แผนที่ระยะทางทางพันธุกรรมจากความถี่รัสเซียโดยเฉลี่ยโดยใช้เครื่องหมายแบบคลาสสิกแสดงระดับความคล้ายคลึงกันของประชากรแต่ละกลุ่มในยุโรปตะวันออกกับกลุ่มยีนรัสเซีย พื้นที่แสงที่ใกล้เคียงที่สุดกับความถี่ยีนรัสเซียกลางตรงบริเวณโซนกลางของยุโรปตะวันออก - จากเบลารุสถึงโวลก้ากลาง (รูปที่ 8.3.4.) โทนสีเข้มเป็นบริเวณที่ห่างไกลจากรัสเซียทางพันธุกรรม มีค่อนข้างน้อย - เรียงตามระดับระยะห่างจากรัสเซียกลาง - เหล่านี้คือภูมิภาคไครเมียและทะเลดำ, แม่น้ำโวลก้าตอนล่าง, รัฐบอลติก, ทางเหนือของรัสเซีย, เฟนนอสกันเดียและเทือกเขาอูราลที่ห่างไกลจากพันธุกรรม
ช่วงของเบลารุสและยูเครนมีความคล้ายคลึงกันกับกลุ่มยีนรัสเซีย ที่น่าแปลกใจคือความแตกต่างทางพันธุกรรมที่เด่นชัดระหว่างรัสเซียเหนือและตะวันออกเฉียงเหนือของยุโรปโดยทั่วไป รวมถึง Vyatka อาณานิคมโนฟโกรอดโบราณ
แน่นอน ประชากรรัสเซียพื้นเมืองซึ่งขณะนี้อาศัยอยู่ในดินแดนเหล่านี้มีคุณลักษณะของประชากรที่หลอมรวมในระดับสูงสุด อย่างไรก็ตาม ไม่น่าเชื่อว่าการมีส่วนร่วมของประชากร Finno-Ugric ที่นี่สูงกว่าในประชากรของ Mordovians และ Chuvash ซึ่งบนแผนที่รวมอยู่ใน "ภูมิภาคทางพันธุกรรมของรัสเซียตอนกลาง" อย่างสมบูรณ์ ความแตกต่างดังกล่าวมีแหล่งที่มาที่เป็นไปได้สามประการ ประการแรก ชั้นใต้ดินของ Finno-Ugric อาจดึงดูดผู้คนที่พูดภาษาฟินนิกตะวันตก และไม่หันไปทางตะวันออก
ประการที่สอง ตามที่ข้อมูลทางโบราณคดีระบุ [Sedov, 1999] การล่าอาณานิคมของโนฟโกรอดมีแหล่งที่มาของชนเผ่าสลาฟที่แตกต่างกัน ซึ่งหมายความว่าไม่เพียง แต่ชั้นล่างเท่านั้น แต่ยังรวมถึง superstratum สลาฟด้วยเช่นกันในภาคเหนือของรัสเซีย ประการที่สาม ในประชากรทางตอนเหนือที่มีขนาดเล็ก ปัจจัยการเบี่ยงเบนทางพันธุกรรมมีพลังมากกว่า ซึ่งสามารถ "นำ" พวกมันออกจากแผ่นดินใหญ่ของรัสเซียได้เช่นกัน เป็นไปได้มากว่าปัจจัยทั้งสามทำพร้อมกัน แต่งานของการวิจัยในอนาคตคือการค้นหาความสัมพันธ์ที่แท้จริงของพวกเขา ที่นี้ เครื่องหมาย unparental สามารถช่วยได้มาก ซึ่งช่วยแยกความแตกต่างของกระแสการอพยพในอวกาศและเวลา
ความใกล้เคียงกับความถี่ "รัสเซียกลาง" จะแสดงโดยส่วนที่มีความหลากหลายมากที่สุดของช่วงรัสเซียรวมถึงส่วนที่ตรงกันข้ามสุดขั้วขององค์ประกอบหลักของกลุ่มยีนยุโรปตะวันออก (ส่วนที่ 8.2.) ภาพที่คล้ายกันสามารถอธิบายได้บนพื้นฐานของสมมติฐานที่ว่าความถี่ "รัสเซียกลาง" นั้นเป็น "ยุโรปกลาง" โดยพื้นฐานแล้วและกลุ่มยีนของรัสเซียนั้นเกิดจากส่วนผสมของส่วนประกอบต่างๆในยุโรปตะวันออก (Finno-Ugric, Slavic, Baltic, เป็นต้น) สมมติฐานนี้ยังได้รับการยืนยันในแผนที่ของระยะทางทางพันธุกรรมจากชาวยูเครน เบลารุส และรัสเซีย ตามตัวบ่งชี้ดีเอ็นเอที่ให้ข้อมูลมากที่สุด - แฮปโลกรุ๊ป Y โครโมโซม
จากประชากรรัสเซีย (เครื่องหมาย DNA อัตโนมัติ)
เช่นเดียวกับข้อมูลจากเครื่องหมายคลาสสิก (รูปที่ 8.3.4) ประชากรของรัสเซียตอนกลางก็ใกล้เคียงกับความถี่ยีนรัสเซียโดยเฉลี่ยอีกครั้ง (รูปที่ 8.3.5.) ชาวเบลารุสซึ่งแทบจะแยกไม่ออกจากลักษณะทั่วไปของรัสเซียในแง่ของความถี่ของเครื่องหมายคลาสสิก และแสดงความแตกต่างเล็กน้อยในแง่ของข้อมูลดีเอ็นเอ ประชากรของเทือกเขาอูราล, คอเคซัส, ภูมิภาคโวลก้าและทางเหนือของรัสเซียค่อนข้างแตกต่างจากความถี่เฉลี่ยของรัสเซีย ดังนั้น ในประเด็นหลักทั้งหมด การใช้ DNA และเครื่องหมายแบบคลาสสิกจึงนำไปสู่ผลลัพธ์ที่คล้ายคลึงกัน ในความเห็นของเรา ความแตกต่างระหว่างแผนที่ทั้งสองนั้นส่วนใหญ่เกิดจากระดับความรู้ของเครื่องหมายประเภทต่างๆ และสามารถคาดหวังได้ว่าเมื่อมีการสะสมข้อมูลเกี่ยวกับความหลากหลายทางดีเอ็นเอ รูปแบบของความแปรปรวนของพวกมันจะเข้าใกล้ผลลัพธ์ที่ได้รับมากขึ้น โดยใช้เครื่องหมายคลาสสิก
ระยะห่างทางพันธุกรรมเฉลี่ยของประชากรยุโรปตะวันออกจากความถี่รัสเซียมีน้อย (d=0.28) ซึ่งอาจเป็นผลมาจากปฏิสัมพันธ์ระยะยาวของแหล่งรวมยีนรัสเซียกับสิ่งแวดล้อม โปรดจำไว้ว่าระยะทางจากชนชาติอินโด - ยูโรเปียนโดยรวมนั้นมีค่าเฉลี่ยเท่ากัน (d=0.28) เมื่อเปรียบเทียบแผนที่เหล่านี้ (รูปที่ 8.3.1 และ 8.3.5) จะมีความคล้ายคลึงกันอย่างเห็นได้ชัด นี่เป็นสิ่งที่เข้าใจได้ เนื่องจากชาวรัสเซียก็เป็นชาวอินโด-ยูโรเปียนเช่นกัน และความถี่ในประชากรรัสเซียก็รวมอยู่ในการคำนวณสำหรับชนชาติอินโด-ยูโรเปียนด้วย เป็นเรื่องแปลกที่ความแตกต่างระหว่างประชากรรัสเซียระหว่างแม่น้ำโวลก้าและวัตกาซึ่งทำเครื่องหมายไว้บนแผนที่ระยะทางจากความถี่เฉลี่ยระหว่างชนชาติอินโด - ยูโรเปียนนั้นยังคงอยู่บนแผนที่ระยะทางจากความถี่รัสเซียโดยเฉลี่ย
ดังนั้น กลุ่มยีนของรัสเซียจึงมีความเกี่ยวข้องอย่างใกล้ชิดกับกลุ่มยีนของชาวยุโรปตะวันออกจำนวนมาก - ในแง่ของความถี่ของยีน เบลารุส ยูเครน มอร์โดเวีย และประชากรยุโรปตะวันออกอื่น ๆ อีกมากมายมีความใกล้ชิดกับรัสเซียมาก เมื่อเราเข้าใกล้คอเคซัสและเทือกเขาอูราลเท่านั้นที่กลุ่มยีนของประชากรจะแตกต่างไปจากลักษณะทั่วไปของยีนรัสเซียอย่างชัดเจน ผลลัพธ์นี้ไม่ใช่เรื่องที่ไม่คาดคิด เนื่องจากการตั้งถิ่นฐานของรัสเซียเหนือดินแดนอันกว้างใหญ่และการแลกเปลี่ยนยีนอย่างเข้มข้นนอกขอบเขต "ดั้งเดิม" กับชนชาติโดยรอบนั้นชัดเจน ค่อนข้างน่าสนใจที่แผนที่ยีนภูมิศาสตร์แสดงให้เห็นการมีอยู่ของกำแพงภูเขาสองแห่ง (คอเคซัสและเทือกเขาอูราล) ซึ่งจำกัดการขยายตัวเชิงพื้นที่ของแหล่งรวมยีน
จากประชากรรัสเซีย (เครื่องหมายโครโมโซม Y)
การ์ดใบนี้มีคุณสมบัติสองประการ ประการแรก เราเห็นทั้งยุโรป ไม่ใช่แค่ครึ่งทางตะวันออก ประการที่สอง ความสามารถในการแยกความแตกต่างของเครื่องหมายโครโมโซม Y นั้นสูงกว่ามาก ดังนั้นความแตกต่างระหว่างประชากรรัสเซียและเพื่อนบ้านจึงเด่นชัดกว่า แม้จะมีช่วงระยะห่างที่ "กว้างกว่า" แต่ช่วงเวลาของระยะทางสูงสุดก็ครอบงำแผนที่ - ตามเครื่องหมายของโครโมโซม Y เกือบทั้งหมดของยุโรปจะแตกต่างอย่างมากจากแหล่งรวมยีนของรัสเซีย (รูปที่ 8.3.6) . เฉพาะประชากรรัสเซียและเบลารุสเท่านั้นที่ใกล้เคียงที่สุดกับความถี่รัสเซียโดยเฉลี่ย ระดับความใกล้เคียงโดยเฉลี่ยจะแสดงโดย Ukrainians ชนชาติสลาฟตะวันตก (โปแลนด์ เช็ก สโลวัก) และประชาชนในภูมิภาคโวลก้า เช่นเดียวกับในแผนที่ก่อนหน้านี้ ประชากรรัสเซียตอนเหนือแสดงอัตลักษณ์ที่เด่นชัด แตกต่างอย่างมากจากกลุ่มยีนรัสเซียโดยเฉลี่ย
เราเห็นว่าเครื่องหมายของโครโมโซม Y ยืนยันรูปแบบที่ระบุไว้ก่อนหน้านี้ของความคล้ายคลึงกันของกลุ่มยีน "รัสเซียกลาง" กับชนชาติสลาฟตะวันออกอื่น ๆ และประชาชนในภูมิภาคโวลก้าและความแตกต่างของรัสเซียเหนือ เนื้อหาข้อมูลสูงของโครโมโซม Y ทำให้รูปแบบเหล่านี้นูนขึ้นมากกว่าเครื่องหมายประเภทอื่น และการพิจารณาจากขนาดของทั้งยุโรปทำให้ Yeshe และ Poles อยู่ในรายชื่อบุคคลที่คล้ายกับกลุ่มยีนรัสเซีย
จากเบลารุส (เครื่องหมายคลาสสิก)
ในแผนที่ก่อนหน้านี้ (รูปที่ 8.3.4., 8.3.5., 8.3.6.) เราพบว่าประชากรจำนวนมากในยุโรปตะวันออกมีความคล้ายคลึงกับกลุ่มยีนของรัสเซีย
สิ่งสำคัญคือต้องเข้าใจ: ประชากรทั้งหมดเหล่านี้อยู่ใกล้กับกลุ่มยีนรัสเซียหรือกับประชากรสลาฟตะวันออกที่หลากหลายหรือไม่? กล่าวอีกนัยหนึ่ง: ความลับของความคล้ายคลึงกันนี้อยู่ในประวัติศาสตร์ชาติพันธุ์ของชาวรัสเซียหรือในการขยายตัวของชาวสลาฟตะวันออกโดยรวมและอาจอยู่ใน "ดั้งเดิม" ก่อนการขยายตัว ความคล้ายคลึงกันของชาวสลาฟและฟินโน- กลุ่มยีน Ugric?
เพื่อตอบคำถามนี้ เราได้วิเคราะห์ความใกล้ชิดของยีนพูลยุโรปตะวันออกกับชาวเบลารุส ซึ่งเป็นกลุ่มชาติพันธุ์สลาฟตะวันออกอีกกลุ่มหนึ่ง ซึ่งใกล้ชิดกันมากในด้านภูมิศาสตร์ ชาติพันธุ์วิทยา และประเภทมานุษยวิทยาสำหรับชาวรัสเซีย
ในรูป 8.3.7. แผนที่ระยะทางทางพันธุกรรมของประชากรของยุโรปตะวันออกจากความถี่ยีนเฉลี่ยของเบลารุสสำหรับเครื่องหมายยีนคลาสสิกชุดใหญ่ - ให้ 57 อัลลีลจาก 21 loci เราเห็นภาพที่ชัดเจนโดยพื้นฐานแล้วแตกต่างจากธรรมชาติของความแปรปรวนของยีนรัสเซีย เกือบทุกภูมิภาคซึ่งมีประชากรอยู่ใกล้กับกลุ่มยีนเบลารุสมากที่สุดตั้งอยู่ในอาณาเขตของเบลารุส นอกขอบเขตของเบลารุส ระยะทางทางพันธุกรรมเพิ่มขึ้นอย่างรวดเร็วจนถึงค่าที่มีนัยสำคัญ ซึ่งบ่งชี้ถึงความแตกต่างทางพันธุกรรมที่ชัดเจนระหว่างกลุ่มยีนของชาวเบลารุสและกลุ่มยีนในยุโรปตะวันออกโดยรวม
แผนที่จะรวบรวมความคิดริเริ่มทางพันธุกรรมของกลุ่มยีนเบลารุส ซึ่งบ่งชี้ถึงความไวสูงของวิธีการวัดระยะทางพันธุกรรม ควรสังเกตว่าความแตกต่างที่ชัดเจนระหว่างกลุ่มยีนเบลารุสและกลุ่มยีนของดินแดนใกล้เคียงเป็นผลที่ไม่คาดคิดที่สำคัญ เนื่องจากข้อมูลทางมานุษยวิทยามักจะล้มเหลวในการเปิดเผยความแตกต่างที่เด่นชัดระหว่างชาวเบลารุสกับกลุ่มเพื่อนบ้าน [Alekseeva, 1973; เดอยาบิน, 1999]. แน่นอนว่าความคิดริเริ่มทางพันธุกรรมของชาวเบลารุสมีความเกี่ยวข้องกันมาก: ปรากฏเฉพาะในระดับเบลารุสราวกับว่าผ่านกล้องจุลทรรศน์ด้วยความละเอียดขนาดใหญ่ของแผนที่เพื่อดูรายละเอียดที่ละเอียด จำได้ว่าในระดับที่แตกต่างกัน - บนแผนที่ของระยะทางทางพันธุกรรมจากรัสเซีย - ชาวเบลารุสแทบจะแยกไม่ออกจากรัสเซียในรัสเซียตอนกลาง ไม่ว่าในกรณีใด ชาวเบลารุสก็เหมือนพวกเขามากกว่าประชากรรัสเซียในรัสเซียเหนือเอง
ดังนั้น ไม่เหมือนกับกลุ่มของรัสเซีย กลุ่มยีนของเบลารุสไม่ได้อยู่ใกล้กับกลุ่มยีนยุโรปตะวันออกโดยรวม ดังนั้นความคล้ายคลึงทางพันธุกรรมในระดับสูงของประชากรรัสเซียกับประชากรในดินแดนยุโรปตะวันออกส่วนใหญ่จึงไม่ใช่ลักษณะทั่วไปของชนชาติสลาฟตะวันออกทั้งหมด แต่เป็นลักษณะของกลุ่มยีนรัสเซีย
จากเบลารุส (Y CHROMOSOME MARKERS)
ข้อสรุปนี้ยังได้รับการยืนยันโดยข้อมูลเกี่ยวกับโครโมโซม Y แผนที่ระยะทางจากชาวเบลารุส (รูปที่ 8.3.8.) สร้างขึ้นในช่วงเวลาเดียวกันกับชาวรัสเซีย (รูปที่ 8.3.6.) แต่โซนทางพันธุกรรมที่คล้ายกับกลุ่มยีนเบลารุสนั้นเล็กกว่าอย่างเห็นได้ชัด: รวมเฉพาะชนชาติสลาฟ (ทั้งชาวสลาฟตะวันออกยกเว้นยูเครนตะวันตกและประชากรสลาฟตะวันตก) แต่ไม่รวมผู้คนในภูมิภาคโวลก้าและอูราล ดังนั้นความคล้ายคลึงทางพันธุกรรมกับประชากรที่ไม่ได้พูดภาษาสลาฟในยุโรปตะวันออกจึงเป็น "อภิสิทธิ์" ของแหล่งรวมยีนรัสเซีย ตรงกันข้ามกับกลุ่มยีนของเบลารุส ซึ่งแตกต่างอย่างมากจากชนชาติเหล่านี้ในแม่น้ำโวลก้าและอูราล
จากชาวยูเครน (Y CHROMOSOME MARKERS)
เพื่อความสมบูรณ์ของการพิจารณาชนชาติสลาฟตะวันออก เรายังนำเสนอแผนที่ระยะทางจากยูเครน (รูปที่ 8.3.9.) เป็นการเตือนความจำมากของแผนที่จากชาวเบลารุสที่เพิ่งตรวจสอบ เฉพาะโซนที่ใกล้เคียงที่สุดเท่านั้นที่จะถูกย้ายไปยังพื้นที่ของ Ukrainians เองและโซนนี้ยังรวมถึงประชากรรัสเซียตอนใต้และเบลารุสด้วย และชนชาติที่ไม่ใช่สลาฟในยุโรปตะวันออก ซึ่งค่อนข้างใกล้เคียงกับประชากรรัสเซีย อยู่ห่างไกลจากแหล่งรวมยีนของยูเครน เนื่องจากพวกเขามาจากแหล่งรวมยีนของเบลารุส สิ่งนี้ยืนยันความถูกต้องของการตีความของเราว่าการล่าอาณานิคมของสลาฟของที่ราบยุโรปตะวันออกพร้อมด้วยการดูดซึมของประชากร Finno-Ugric ซึ่งส่วนใหญ่เกี่ยวข้องกับบรรพบุรุษของประชากรรัสเซียสมัยใหม่จากเทือกเขาสลาฟทั้งหมด






เป็นครั้งแรกที่นักวิจัยได้สร้างแผนที่โดยละเอียดของโครงสร้างทางพันธุกรรมของประชากรในสหราชอาณาจักร ปรากฎว่าประชากรของดินแดนเซลติกไม่ได้เป็นเนื้อเดียวกันทางพันธุกรรมเลยและกลุ่มยีนที่เก่าแก่ที่สุดได้รับการเก็บรักษาไว้ในเวลส์ การเปรียบเทียบ DNA ของชาวอังกฤษและชาวยุโรปในทวีปยุโรปบ่งชี้รายละเอียดทางพันธุกรรมของบรรพบุรุษในแต่ละภูมิภาค ในแหล่งรวมยีนของประชากรสมัยใหม่ เป็นไปได้ที่จะเห็นภาพสะท้อนของเหตุการณ์ที่สำคัญที่สุดในประวัติศาสตร์ของการตั้งถิ่นฐานของเกาะอังกฤษ
การศึกษาความแตกต่างทางพันธุกรรมระหว่างประชากรสมัยใหม่ช่วยให้มองลึกลงไปในประวัติศาสตร์และเห็นร่องรอยของการอพยพของประชากรด้วยการสร้างแหล่งรวมยีนที่ทันสมัย งานนี้ดำเนินการโดยทีมนักวิจัยนานาชาติที่นำโดยผู้เชี่ยวชาญชาวอังกฤษจาก University of Oxford และ University College London บทความของพวกเขาได้รับการตีพิมพ์ในวารสาร Nature เมื่อวันที่ 19 มีนาคม Walter Bodmer, Peter Donnelly และเพื่อนร่วมงานวิเคราะห์รายละเอียดโครงสร้างทางพันธุกรรมของประชากรสหราชอาณาจักร พวกเขาไม่เพียงแต่สร้างแผนที่ทางพันธุกรรมที่แม่นยำมากของประชากรอังกฤษยุคใหม่ แต่ยังพบร่องรอยของการอพยพทางประวัติศาสตร์ที่สำคัญที่สุดและชี้แจงรูปแบบการตั้งถิ่นฐานในเกาะอังกฤษ
กลุ่มยีนสะท้อนถึงภูมิศาสตร์
นักวิจัยได้รวบรวม DNA จาก 2039 คนจากพื้นที่ต่าง ๆ ของสหราชอาณาจักร เมื่อเลือกพวกเขาได้รับคำแนะนำจากกฎต่อไปนี้: บรรพบุรุษทั้งหมดของบุคคลในรุ่นที่สาม (ทั้งคุณย่าและปู่ทั้งสอง) ต้องอยู่ห่างจากกันไม่เกิน 80 กม. เงื่อนไขนี้ทำให้ได้รับ "ภาพรวม" ของโครงสร้างทางพันธุกรรมของประชากรชาวอังกฤษในช่วงปลายศตวรรษที่ 19 และต้นศตวรรษที่ 20
ในการเปรียบเทียบ DNA นักวิทยาศาสตร์ได้ใช้ single nucleotide polymorphisms (SNPs) มากกว่า 500,000 ตัวที่กระจัดกระจายไปทั่วจีโนม การวิเคราะห์ดำเนินการโดยใช้วิธีทางสถิติ fineSTRUCTURE และ GLOBETROTTER วิธีการเหล่านี้จัดกลุ่มตัวอย่าง DNA ตามความคล้ายคลึงกัน โดยไม่คำนึงว่าพวกเขาจะเก็บตัวอย่างที่ใดในสหราชอาณาจักร และโดยการวางกลุ่มตัวอย่างแต่ละคนไว้ตรงกลางระหว่างบ้านเกิดของปู่ย่าตายาย พวกเขาสามารถเห็นได้ว่าความคล้ายคลึงทางพันธุกรรมมีความสัมพันธ์กับที่ตั้งทางภูมิศาสตร์อย่างไร
กลุ่มพันธุกรรมที่จัดกลุ่มตัวอย่าง DNA 2039 จากประชากรในสหราชอาณาจักร
เป็นเรื่องบังเอิญที่อัศจรรย์ใจ ดังที่เห็นในภาพ มีการกระจายกลุ่มพันธุกรรม 17 กลุ่มบนแผนที่ตามตำแหน่งทางภูมิศาสตร์ของตัวอย่างที่นำมา กระจุกที่ต่างกันในทางปฏิบัติจะไม่ทับซ้อนกัน และเนื่องจากกลุ่มเหล่านี้ก่อตัวขึ้นจากตัวอย่างทั้งหมด โดยไม่คำนึงถึงภูมิศาสตร์ของพวกมัน การติดต่อนี้สะท้อนถึงความหลากหลายทางพันธุกรรมที่แท้จริงของประชากร ผู้เขียนจึงเน้นย้ำ
ประชากรของเคาน์ตีออร์กนีย์ในหมู่เกาะออร์กนีย์ทางเหนือของสกอตแลนด์กลายเป็นความหลากหลายทางพันธุกรรมมากที่สุด โดยประกอบด้วยสามกลุ่ม ความแตกต่างทางพันธุกรรมสามารถมองเห็นได้ชัดเจนในพื้นที่ต่างๆ ของสหราชอาณาจักร: ทางตอนเหนือของอังกฤษ สกอตแลนด์ และไอร์แลนด์เหนือ มีความแตกต่างจากทางตอนใต้ของอังกฤษ เวลส์ตอนเหนือแตกต่างจากทางใต้ และคอร์นวอลล์แยกเป็นกระจุก ขอบเขตของกระจุกดาวมักจะตรงกับเขตแดนสมัยใหม่ของจังหวัดทางประวัติศาสตร์ เช่น ที่ชายแดนคอร์นวอลล์และเดวอน ที่ชายแดนอังกฤษและเวลส์ กลุ่มที่ใหญ่ที่สุด (แสดงเป็นสี่เหลี่ยมสีแดงบนแผนที่) ครอบคลุมพื้นที่ส่วนใหญ่ของอังกฤษตอนกลางและตอนใต้ และชายฝั่งตะวันออก รวมตัวอย่าง DNA เกือบครึ่งหนึ่งที่รวมอยู่ในการศึกษา
ผู้เขียนสร้างต้นไม้สายวิวัฒนาการของประชากรอังกฤษ แสดงให้เห็นว่ากลุ่มเหล่านี้ก่อตัวอย่างไร แยกกิ่งก้านหลักอย่างไร อย่างที่คุณเห็นบนต้นไม้ ในระยะแรกสุด กิ่งก้านของประชากรของหมู่เกาะออร์คนีย์แยกตัวออกจากกัน ในขั้นต่อไป เวลส์ก็แยกตัว ทางแยกที่ตามมาบนต้นไม้นำไปสู่การแยกทางเหนือและใต้ของเวลส์ การแยกทางเหนือของอังกฤษ สกอตแลนด์ และไอร์แลนด์เหนือออกจากส่วนอื่นๆ ของอังกฤษ และการแยกคอร์นวอลล์ออกจากกลุ่มที่เหลือ
ดังนั้นแนวคิดเรื่องความสม่ำเสมอทางพันธุกรรมของประชากรเซลติกของบริเตนใหญ่ในฐานะที่เก่าแก่ที่สุดซึ่งไม่ได้รับอิทธิพลจากการอพยพของชาวแซกซอนจึงไม่ได้รับการยืนยัน ในทางตรงกันข้าม ส่วนเซลติกของบริเตนใหญ่ (สกอตแลนด์ ไอร์แลนด์เหนือ เวลส์ และคอร์นวอลล์) กลับกลายเป็นว่ามีความหลากหลายทางพันธุกรรมมากกว่าพื้นที่ที่เหลือ
แม้ว่ากระจุกในประชากรอังกฤษจะมีความโดดเด่นอย่างชัดเจน เมื่อนักวิทยาศาสตร์วัดระยะห่างทางพันธุกรรมระหว่างพวกเขา มันก็มีขนาดเล็ก (เฉลี่ย 0.002 สูงสุด 0.007) ซึ่งหมายความว่าความแตกต่างที่สังเกตได้นั้นละเอียดอ่อนมากและสามารถมองเห็นได้ด้วยการ "ซูม" ที่ชัดเจนเท่านั้น ซึ่งเป็นสิ่งที่นักวิจัยสร้างขึ้นโดยใช้วิธีการทางสถิติที่ทรงพลัง สำหรับการเปรียบเทียบ พวกเขาวิเคราะห์ข้อมูลโดยใช้วิธี Principal Component Analysis (PCA) และ ADMIXTURE มาตรฐาน ทั้งสองวิธีแสดงให้เห็นการแยกสาขาของออร์กนีย์และเวลส์ แต่ไม่ได้เปิดเผยโครงสร้างทางพันธุกรรมที่ละเอียดกว่าของประชากร
รอยเท้าทวีปในจีโนมอังกฤษ
ความแตกต่างทางพันธุกรรมภายในประชากรชาวอังกฤษอาจเป็นผลมาจากการแยกกลุ่มประชากรบางกลุ่มออกจากกัน แต่ยังสะท้อนถึงการมีส่วนร่วมของการอพยพต่าง ๆ และกระบวนการผสมผู้อพยพกับประชากรพื้นเมือง เพื่อจัดการกับกรณีหลัง นักวิจัยใช้วิธีการเดียวกันเพื่อเปรียบเทียบตัวอย่าง DNA จากประชากรอังกฤษกับตัวอย่างจากกว่า 6,000 ทวีปยุโรปจาก 10 ประเทศ ตัวอย่างเหล่านี้ถูกแบ่งออกเป็น 51 กลุ่มตามความคล้ายคลึงทางพันธุกรรม

โปรไฟล์ทางพันธุกรรมของยุโรปสำหรับ 17 กลุ่มของอังกฤษ
สำหรับแต่ละกลุ่มของอังกฤษ นักวิทยาศาสตร์ประเมินการมีส่วนร่วมของแต่ละกลุ่มทวีปและกำหนดรายละเอียดทางพันธุกรรมของบรรพบุรุษ การวิเคราะห์พบว่ากลุ่มยุโรปบางกลุ่ม เช่น จากเยอรมนีตะวันตก ส่วนเฟลมิชของเบลเยียม ฝรั่งเศสตะวันตกเฉียงเหนือ ฝรั่งเศสตอนใต้ และสเปน มีส่วนสำคัญต่อการกำเนิดของกลุ่มอังกฤษเกือบทั้งหมด บางคนมีส่วนสนับสนุนเฉพาะบางกลุ่มเท่านั้น ตัวอย่างเช่น ชาวนอร์เวย์มีผลกระทบอย่างมีนัยสำคัญต่อการก่อตัวของกระจุกในออร์กนีย์ และน้อยกว่าในสกอตแลนด์และไอร์แลนด์เหนือ
กลุ่มยีนสะท้อนประวัติศาสตร์
ข้อมูลทางพันธุกรรมสะท้อนให้เห็นร่องรอยของการอพยพไปยังเกาะอังกฤษในช่วง 10,000 ปีที่ผ่านมา ตามธรรมชาติของอิทธิพลของกลุ่มทวีปที่มีต่อกระจุกของอังกฤษ นักวิจัยแยกแยะเวลาสัมพัทธ์ของเหตุการณ์การย้ายถิ่นฐาน กลุ่มที่พบว่ามีส่วนสนับสนุนในทุกกลุ่มชี้ให้เห็นถึงการอพยพในสมัยโบราณ เวลาผ่านไปพอสมควรแล้วสำหรับฮาโพลไทป์ของพวกเขาที่จะแพร่กระจายไปยังส่วนอื่น ๆ ของสหราชอาณาจักร รอยเท้าของทวีปในกลุ่มที่แยกจากกันนั้นเป็นผลมาจากการย้ายถิ่นครั้งล่าสุดอย่างเห็นได้ชัด

เหตุการณ์หลักของการตั้งถิ่นฐานของเกาะอังกฤษ
การพิชิตอังกฤษโดยแองโกล-แซกซอนในศตวรรษที่ 5 ทิ้งร่องรอยที่เห็นได้ชัดเจนที่สุด การมีส่วนร่วมทางพันธุกรรมของการย้ายถิ่นของแองโกล-แซกซอนมีความสำคัญในภาคตะวันออกเฉียงใต้ ภาคกลาง และตอนใต้ของอังกฤษ แม้ว่าจะไม่เกิน 50% ในทุกที่ และในพื้นที่ส่วนใหญ่มีตั้งแต่ 10 ถึง 40% สิ่งนี้บ่งชี้ว่าแองโกล-แอกซอนไม่ได้แทนที่ประชากรในท้องถิ่นของชาวอังกฤษ (เซลติกส์) อย่างสมบูรณ์ แต่ผสมกับมัน กระจุกบางกลุ่มสอดคล้องกับภูมิประเทศของอาณาจักรและกลุ่มชนของแองโกล-แซกซอน ดูเหมือนว่าอาณาเขตเหล่านี้ยังคงรักษาเอกลักษณ์ทางชาติพันธุ์และพันธุกรรมไว้เป็นเวลาหลายศตวรรษ
ในประชากรของหมู่เกาะออร์คนีย์ 25% ของยีนพูลมีสาเหตุมาจากบรรพบุรุษของแหล่งกำเนิดของนอร์เวย์ ซึ่งเป็นร่องรอยของการรุกรานหมู่เกาะไวกิ้งของนอร์เวย์ซึ่งเกิดขึ้นในศตวรรษที่ 9 ข้อมูลทางพันธุกรรมยังระบุด้วยว่าชาวไวกิ้งนอร์เวย์ไม่ได้แทนที่ประชากรพื้นเมือง แต่ผสมกับมัน แต่ไม่พบร่องรอยทางพันธุกรรมที่ชัดเจนของพวกไวกิ้งเดนมาร์กซึ่งในขณะเดียวกันก็ควบคุมส่วนสำคัญของอังกฤษไม่ได้
สำหรับการตั้งถิ่นฐานที่เก่าแก่ที่สุดของเกาะอังกฤษ - หลังจากสิ้นสุดการเยือกแข็งครั้งสุดท้าย - ร่องรอยของมันถูกเก็บรักษาไว้อย่างดีที่สุดในประชากรของเวลส์ หลักฐานยังชี้ให้เห็นถึงการอพยพครั้งสำคัญที่ตามมาจากทวีปนี้ แม้กระทั่งก่อนการพิชิตบริเตนโดยจักรวรรดิโรมันในตอนต้นของสหัสวรรษแรก อิทธิพลทางพันธุกรรมของการย้ายถิ่นนี้แพร่กระจายไปทั่วอังกฤษ สกอตแลนด์ และไอร์แลนด์เหนือ แต่เกือบจะข้ามเวลส์ไป ซึ่งมีแหล่งรวมยีนที่เก่าแก่ที่สุด
ผู้เขียนศึกษาแสดงให้เห็นอย่างน่าเชื่อถือว่าพันธุศาสตร์สามารถเสริมและปรับแต่งข้อมูลทางโบราณคดีและภาษาศาสตร์เพื่อสร้างประวัติศาสตร์ของประชากรได้อย่างไร การเลือกข้อมูลเบื้องต้นและวิธีการทางสถิติที่มีประสิทธิภาพช่วยให้พวกเขาได้รับแผนที่พันธุกรรมโดยละเอียดของประเทศและมองย้อนกลับไปในอดีตของประชากรอังกฤษ
แหล่งที่มา:
โครงสร้างทางพันธุกรรมที่ละเอียดของประชากรอังกฤษ
Stephen Leslie, Bruce Winney, Garrett Hellenthal, Dan Davison, Abdelhamid Boumertit, Tammy Day, Katarzyna Hutnik, Ellen C. Royrvik, Barry Cunliffe, Wellcome Trust Case Control Consortium, International Multiple Sclerosis Genetics Consortium, Daniel J. Lawson, Daniel Falush, Colin ฟรีแมน, แมตติ พิริเนน, ไซม่อน ไมเยอร์ส, มาร์ค โรบินสัน, ปีเตอร์ ดอนเนลลี & วอลเตอร์ บอดเมอร์
Nature, 2015, v.519, 7543, ดอย:10.1038/nature14230
http://www.nature.com/nature/journal/v519/n7543/full/nature14230.html