Interaktivt genetisk kort over verden. Genetik af russiske ukrainere, hviderussere og tatarer, slaver og kaukasiere, jøder, finner og andre befolkningsgrupper
I denne undersøgelse bruges mere end 80.000 autosomale snips og referencepunkter - dette er en meget høj opløsning, der gør det muligt at fange selv relativt små påvirkninger på det genetiske niveau i hovedparten af mennesker. De sammenlignende analysedata blev taget fra en åben undersøgelse af V. Verenich, en specialist i sammenlignende analyse af genetiske komponenter. Selve de genetiske regnemaskiner er placeret på GedMatch-tjenesten og giver enhver mulighed for at finde ud af deres komparative position på den genetiske graf. For at gøre dette er det nok at have resultaterne af en autosomal test fra FTDNA eller 23andMe. I slutningen af undersøgelsen leveres kort over geografisk fordeling og frekvensmaksima for de vigtigste autosomale komponenter fra MDLP World-22-projektet.
Graferne nedenfor viser hovedkomponenterne og deres gennemsnitlige procentdel for hver af populationerne. Én linje er den procentvise opdeling for én population. Hver division (lodret streg) svarer til 10 %, og navnene på de autosomale komponenter er i samme rækkefølge fra venstre mod højre som i forklaringen fra top til bund. Jo mere ens procentdelen af den samlede genetik er blandt forskellige folk, jo mere ens ligner figuren i den givne graf. Så lad os komme i gang...
Genetik af tyskere, litauere, russere, svenskere, finner osv.
Denne graf viser de vigtigste genetiske komponenter for europæiske folk og er afstemt efter faldet i den østeuropæiske komponent (nordøsteuropæisk) i forskellige populationer. Som du kan se, er alle europæiske folkeslag ganske forskellige genetisk, og med genetiske komponenter i deres sæt af samme oprindelse, er de ikke desto mindre i meget forskellige procenter. For alle slaver og baltere generelt er en af de mest betydningsfulde denne del af Østeuropa, som er på sit maksimum for litauere og hviderussere. Sandsynligvis siden tiden for den arkæologiske "Corded Ware-kultur" har disse landes territorium været centrum for oprindelsen af denne komponent. Det er repræsenteret af mere end 80 % af litauerne og kun 20 % af italienerne.Lilla repræsenterer Atlanto-Middelhavskomponenten, og den øges, når du bevæger dig fra nordøst til sydvest. Så blandt finnerne når det et gennemsnit på 15 %, og blandt italienerne 40 %. Resten af komponenterne er mindre udtalte.
Genetik af russiske ukrainere hviderussere

Dette diagram viser østslaverne - Russere, hviderussere, ukrainere... Opmærksomheden henledes på ligheden mellem de genetiske mønstre hos de tre listede folkeslag, og inden for fejlmarginen adskiller de sig ganske ubetydeligt - hos ukrainerne og sydrusserne er der en lille stigning i den vestasiatiske komponent, og i de nordlige russere er der er en lille stigning i en af de sibiriske komponenter, kaldet betinget samojeder, og en stigning i komponenterne i Europas mesolitikum til omkring 10%, hvilket ifølge sidstnævnte indikator bringer dem tættere på den tysktalende befolkning i Skandinavien - svenskere.
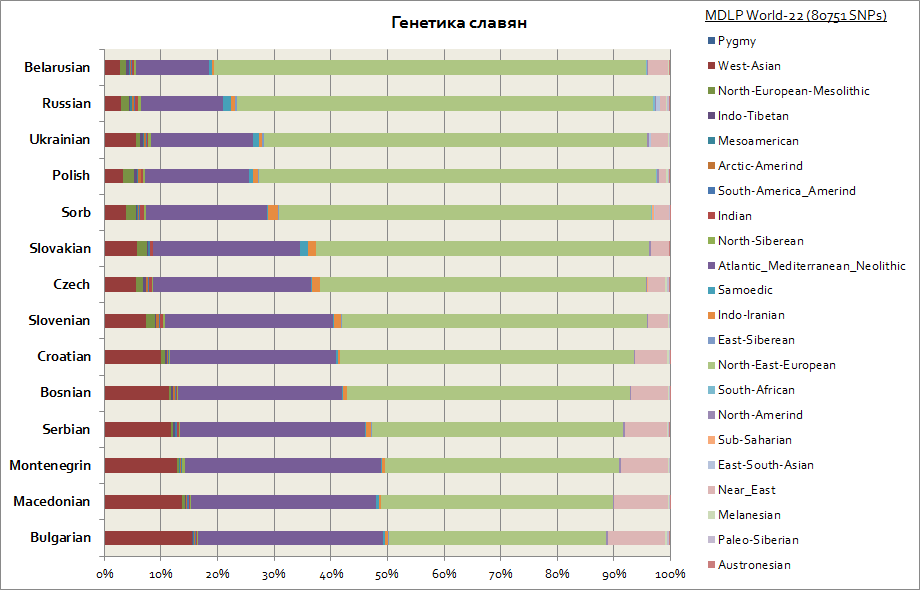
Denne grafik skildrer alle slaverne, inklusive vestlige - polakker og tjekkere, såvel som sydlige - serbere, bulgarere, makedonere osv.
Hovedkomponenterne for alle slaver er 2. Disse er østeuropæiske og Atlanto-Middelhavsområdet. Den første er på maksimum for hviderusserne, og den anden for alle de sydlige slaver - serbere, makedonere, bulgarere. Den østeuropæiske komponent er mere primær blandt slaverne, og Atlanto-Middelhavet er mere erhvervet, da slaverne migrerede til Balkan. Vestukrainere og slovakker har en svag stigning i samojed-komponenten i forhold til naboslaviske folk - hviderussere, tjekkere, polakker; det er sandsynligvis et genetisk spor af den middelalderlige migration af hunnerne og ugrierne til Centraleuropa.
Genetik af slaver, russere og tatarer, tyskere, kaukasiere, jøder osv.
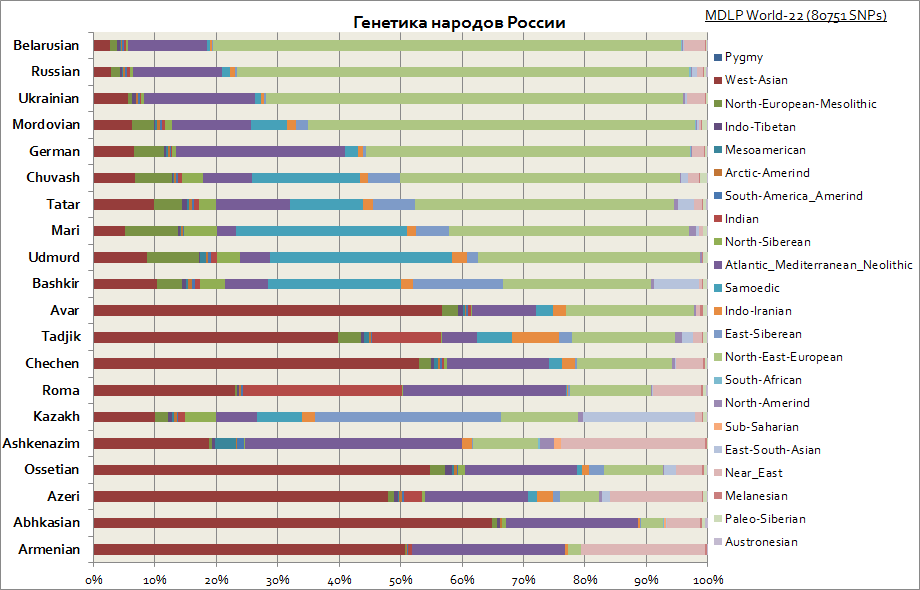
Denne graf viser de forskellige oprindelser blandt folkene i Rusland. Som du kan se, blandt slaverne er den østeuropæiske komponent den vigtigste, og blandt folkene i Volga-regionen er andelen af de sibiriske komponenter stigende. Mens for kaukasierne er den mest karakteristiske den vestasiatiske komponent, Middelhavet og Mellemøsten.
Genetik af finner, ugriere, udmurtere, ungarere, samer osv.

Som du kan se, er finnerne, vepsianerne og karelerne karakteriseret ved en lignende genetisk oprindelse som slaverne. De har også den højeste østeuropæiske komponent, faldende tættere på Ural- og Volga-regionen, med en stigning i de sibiriske komponenter i denne region. Alle finsk-ugriske folk har også en betydelig komponent af Europas mesolithikum, som når næsten 80% blandt samerne og er forbundet med den præ-indoeuropæiske og præ-neolitiske befolkning i Europa. Ungarerne som helhed er karakteriseret ved et sæt af de samme genetiske komponenter som for andre populationer i Karpaterne og Centraleuropa.
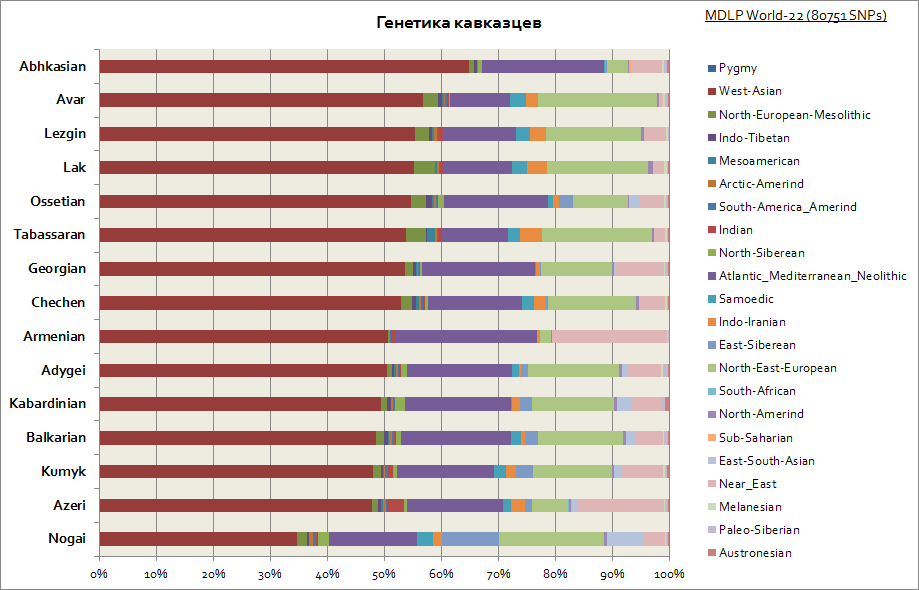
Som du kan se, er hele Kaukasus karakteriseret ved en relativt ens genetisk oprindelse - dette er en stor del af den vestasiatiske komponent og Middelhavet. Kun Nogais er lidt slået ud - deres andel af de sibiriske komponenter stiger.

Som det kan ses, har Ashkenazim og Sephardim en høj frekvens af de vestasiatiske, Atlanto-Middelhavs- og Mellemøstlige komponenter. Samtidig har Ashkenazi en lille stigning i den sibiriske komponent, som sandsynligvis er forbundet med Khazar-arven og væksten på op til 30% af den østeuropæiske komponent, hvilket ifølge denne indikator bringer dem tættere på lande i det sydlige Europa.
Kun etiopiske jøder og indiske jøder er særligt ude af deres "selskab". Førstnævnte har en høj andel af Afrika syd for Sahara (op til 40%), mens sidstnævnte har en høj andel af den sydasiatiske genetiske komponent, konventionelt kaldet indisk (op til 50%).
Genetik af tatarer, bashkirer, aserbajdsjanere, chuvash osv.
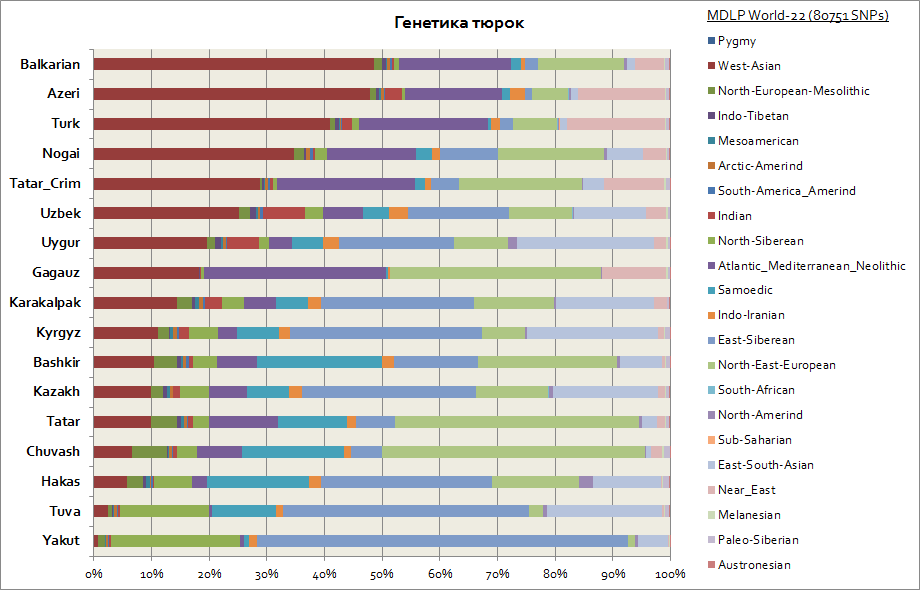
Med hensyn til genetik viste tyrkerne sig at være en af de mest heterogene etniske grupper, derfor adskiller deres genetiske komponenter sig betydeligt. Så i betragtning af at tyrkernes primære hjemland er Sibirien, beholdt sådanne folk som yakuterne, tuvanerne og Khakass den største procentdel af den østsibiriske autosomale komponent, som når fra 30 til 65%. Denne genetiske komponent er også den vigtigste blandt kirgiserne og kasakherne. Resten af komponenterne bringer tyrkerne tættere på folkene fra bopælsregionerne. Så for yakuterne og tuvanerne er disse de nordsibiriske og samodianske komponenter. I alt disse 3 sibiriske komponenter blandt yakuterne er det op til 90%, blandt tuvinerne op til 70%, med en stigning til 20% af den øst-sydasiatiske komponent, som i højere grad er forbundet med migrationsstrømmene for befolkningen i Østasien . For bashkirerne er andelen af 3 sibiriske komponenter op til 45%, og den sydøstasiatiske komponent er op til 10%. Tatarerne har data om 3 sibiriske genetiske komponenter i gennemsnit fra 25 til 50%. Samtidig er andelen af komponenter, der er karakteristiske for den kaukasiske befolkning blandt bashkirerne, op til 45%, og blandt tatarerne i gennemsnit fra 50 til 70%. Aserbajdsjanernes og tyrkernes genetik adskiller sig praktisk talt ikke inden for fejlmarginen; de har ligesom resten af befolkningerne i Kaukasus- og Transkaukasiske regioner en betydelig vestasiatisk komponent (op til 50%) og Atlanterhavet-Middelhavet (op til 20 % i gennemsnit). Andelen af 3 sibiriske komponenter præsenteres af aserbajdsjanere, tyrkere og balkarer - på niveauet 3-7%.
Konklusion
Folks genetik har ikke en direkte og signifikant sammenhæng med fordelingen af sprogfamilier eller med procentdelen af homogene markører - Y-DNA og mt-DNA haplogrupper, repræsenteret i en bestemt population. Den største sammenhæng kan spores efter det territorialgeografiske princip. Således falder andelen af de sibiriske komponenter, der er karakteristiske for den mongoloide race som helhed, gradvist fra øst til vest, og andelen af komponenter, der er karakteristiske for den kaukasiske race, stiger tilsvarende. I grænseområderne langs linjen fra den nordlige del af Ural til Centralasien er deres forhold omtrent det samme. I regionerne øst for Bajkalsøen er de genetiske komponenter, der er karakteristiske for den store kaukasiske race, praktisk talt ikke repræsenteret, samtidig forsvinder i regionerne vest for Pechora-Volga-regionen sibiriske komponenter, der er karakteristiske for den store mongoloide race.Udbredelsen af den østeuropæiske genetiske komponent til Sibirien skete i vid udstrækning allerede i bronzealderen (kultur i Andronov-cirklen), selvom enkelte toppe i det yderste østlige Sibirien blandt tjukchierne kan have været forbundet med russernes migrationer i det 17. århundrede.
Andelen af komponenten syd for Sahara, karakteristisk for den negroide race, er fordelt over hele Afrika - op til det sydlige Middelhav og den nordlige grænse af det afrikanske kontinent, når et maksimum i sin ækvatoriale del, og ud over dets grænser gør det praktisk talt ikke forekomme; lys baggrund almindelig på den arabiske halvø og den sydlige del af det iranske plateau.
Geografi af genetiske komponenter
Alexey Zorrin
Projekt
Genetisk analyse er længe og fast gået ind i antallet af rutineprocedurer inden for sådanne anvendte vidensområder som retsvidenskab og medicin. Forbedringen af metoderne til genkendelse af genetiske markører finder som regel anvendelse i grundlæggende videnskabelige problemer.
For eksempel takket være nye tekniske løsninger, fuldt sekventeret mtDNA forhistorisk mand af neandertaler-typen, og i den nærmeste fremtid håber forskerne at dechifrere dets komplette nukleare genom og sammenligne det med mennesket. Vores egen historie bliver i stigende grad åbenbaret for os ikke gennem arkæologiske fund, men gennem metoderne til genetisk analyse. For eksempel lærte vi den omtrentlige kronologi af Jordens bosættelse og fandt ud af, at på det meget tidlige stadium af dens udvikling var menneskeheden næsten forsvandt fra jordens overflade på grund af tørke.
Data indkodet i flere milliarder nukleotider af DNA kan dog fortælle meget mere om en bestemt person og om menneskeheden som helhed. John Novembre fra University of California i Los Angeles og hans kolleger viste, hvordan en storstilet genetisk analyse af mere end tre tusinde europæere allerede har gjort det muligt at afsløre strukturen af befolkningen i Europa og finde ud af, hvordan genetisk forskellige repræsentanter for små Europæiske stater er.
Nu kan videnskabsmænd med en nøjagtighed på flere hundrede kilometer kun bestemme oprindelsesstedet for enhver europæer på basis af hans DNA.
Der findes mange matematiske metoder til at løse dette ikke helt klart formulerede problem; den mest almindelige inden for genetik er den primære komponentanalyse, som gør det muligt at bestemme sådanne "akser" i et multidimensionelt datasæt, langs hvilke genomforskelle er mest mærkbare. Novembre og hans kolleger udnyttede det og begrænsede sig til to sådanne akser.
Ved at placere alle næsten femten hundrede punkter på flyet, så de et billede, der påfaldende minder om det politiske kort over Europa.
Det portugisiske og spanske genom var på dette kort til venstre for franskmændene, tyskerne - til højre var "italienerne" placeret under franskmændene, og briterne var over dem. Heldigvis kan du vride det resulterende kort, som du vil - det ændrer ikke på de geometriske forhold mellem punkternes positioner. Så forskerne skulle kun sammenligne den genetiske graf med et geografisk kort - den såkaldte klinalitet ifølge den sammensatte parameter svarende til hovedaksen - og fastslå, at den vigtigste "hovedkomponent" svarer til den syd-syd-østlige - nord-nord-vest akse, og nemlig i denne form og præsentere kortet for kolleger.
Det genetiske kort over Europa af John Novembre og hans kolleger. Nederst til højre - et mindre slående resultat af et lignende værk af Manfred Kaiser, udgivet for en måned siden. // John Novembre / M.Kaiser
Selvfølgelig var det endelige kort ikke uden fejl. Ikke nok med det, individuelle genomer har spredt sig langt ud over grænserne for de lande, der svarer til dem. Russerne slog sig ned på denne graf blandt tjekkerne og polakkerne og befandt sig meget "vest" for ukrainerne, mens slovakkerne generelt faldt dybt under alperne og befandt sig helt for enden af den italienske "støvle". Dette kan dog forklares med særegenhederne ved den magre prøve af begge: der var kun seks russere i de oprindelige data, og der var kun en slovakisk.
Ikke desto mindre, ved kun at bruge dette kort, kan 90% af mennesker angive deres hjemland med en nøjagtighed på 700 km og 50% - op til 300 km.
Dette er på trods af, at 200 tusind "bogstaver" af DNA - mindre end 0,01% af hele den menneskelige genetiske kode.
Det ses også meget tydeligt på grafen, at europæernes-sydlændinges genetiske diversitet er meget større end de nordlige folks. Disse data, efter Novembras mening, kan let forklares med historien om Europas bosættelse af mennesker. Efter at Homo sapiens dukkede op her for 35 tusinde år siden, kun 15 tusinde år senere, med gletsjernes tilbagetog, fandt den første massevandring af folk mod nord sted, og efter yderligere 10 tusinde år, med fremkomsten af landbruget, fandt nye grupper af sydlændinge drog mod nord, som alle disse årtusinder repræsenterede imponerende, velformede befolkninger.
I øvrigt lignende Arbejde, baseret på meget af de samme genetiske data, blev offentliggjort i Current Biology for en måned siden. Det er værd at bemærke, at dets leder, Manfred Kaiser, formåede at opnå et lignende, men ikke så saftigt billede ved hjælp af den samme metode til hovedkomponenterne. Denne analyse involverede 2.500 genomer omhyggeligt samlet af GlaxoSmithKline, et genjagtfirma, der er ansvarligt for bivirkningerne af menneskelig stofbrug.

"Genetisk kort" over befolkningen i Schweiz // John Novembre
Samtidig var metoden med hovedkomponenter i begge tilfælde ude af stand til at opdele europæerne i separate genetiske klynger. Med andre ord er europæernes genetiske mangfoldighed så lille, at selv en halv million mononukleotidpolymorfier ikke er nok til kun at opdele tre tusinde mennesker i små undergrupper.
"Vi kan nogle gange godt lide at tale om" russiske gener "," polske gener "eller" tatariske "gener. Men denne artikel viser os tydeligt, at der ikke er sådanne gener!"
- kommenterede disse værker til Gazeta.Ru, en af de vigtigste nationale specialister i genogeografi Oleg Balanovsky fra laboratoriet for menneskelig befolkningsgenetik ved det medicinske genetiske forskningscenter ved det russiske akademi for medicinske videnskaber, Kendt til vores læsere.
"Først efter at have studeret hundrede tusinde gener, var forfatterne i stand til (og så med en fejl på flere hundrede kilometer) at bestemme de undersøgte europæeres sandsynlige oprindelsessteder. Det er umuligt at gøre dette ved at bruge ét gen, eller endda hundrede gener! - siger Balanovsky. - Der er ikke et enkelt gen, der er karakteristisk for befolkningen i et givet land. Kun en kombination af hundredtusindvis af gener er mere eller mindre specifik.
Det betyder, at begrundelsen for dem, der frygter indflydelse på folkets genpulje gennem dets specifikke gener."
”Vi har ventet på sådan en artikel i lang tid. Teknologien til at analysere en million genetiske markører har eksisteret i flere år. Og i denne artikel bliver disse nye teknologier endelig anvendt til at studere genpuljen i Europa, fortsætter videnskabsmanden. - Dette arbejde viser perfekt kontinuiteten i videnskaben: Når alt kommer til alt, da der for 10-15 år siden arbejdede med mitokondrielt DNA (moder-arvelinje) og Y-kromosomet (faderlig arvelinje), skete det første gennembrud også i genpuljen af Europa, og selv konklusionen om vigtigheden af geografien i rækkefølgen af genpuljen blev gjort på samme måde som nu."
Ifølge Balanovsky giver dagens teknologier os mulighed for at opnå hidtil uset pålidelighed og nøjagtighed af resultater: "Hvad genetik næppe kunne skelne i en tæt tåge for fyrre år siden, noget der for ti år siden begyndte at blive oplyst af to kraftige tågelygter (mitokondrielt DNA og Y). -kromosom), begynder vi nu at se det samme i lyset af en million svagere lys (gener, selvom de er mindre informative, er talrige og fordelt i hele genomet).
"Det næste og sidste skridt - fuld, hundrede procent belysning - kan forventes om et par år, når fuld sekventering af hele genomet bliver tilgængelig - ikke en million nukleotider, som det er nu, men alle milliarder, der findes i genomet, ” forudser den russiske specialist.
Efter hans mening er det eneste spørgsmål, der rejser sig, om det er nødvendigt at vente på så detaljerede data. Faktisk, selv nu, "gør vores to kraftige lys (mitokondrielt DNA og Y-kromosom) og en million svagere lys (men fordelt i hele genomet) os i stand til at se strukturen af den menneskelige genpulje og drage både grundlæggende videnskabelige og anvendte konklusioner ."
Forskeren er bekymret over, at antallet af undersøgte gener i genomet indtil videre er meget større end antallet af mennesker, hvor disse gener er blevet undersøgt.
"Kun studiet af genpuljen - hele sættet af gener - giver os mulighed for at studere genetik af et folk for at hjælpe læger med at stille diagnoser, og for alle mennesker - at spore deres folks eller deres egen families historie," konkluderede Oleg Balanovsky.
5. januar 2013 Udskriv
Vi hører konstant, at russerne ikke er et folk svejset af blod, beslægtet med blod, men et konglomerat af mennesker forenet af en fælles kultur og territorium. Alle husker Putins slagord "Der er ingen rene russere!" og "skrab hver russer, du vil helt sikkert finde en tatar."
De siger, at vi er "meget forskellige i blod", "spirede ikke fra den samme rod", men var en smeltedigel for tatarerne, kaukasiske, tyske, finske, buryatiske, mordoviske og andre folk, der nogensinde kom, trådte ind, ankom på vores jord, og vi tog imod dem alle, lukkede dem ind i huset, tog dem med til vores slægtninge.
Dette er blevet nærmest et aksiom i brug blandt politikere, der udvander begrebet russisk, og samtidig er det for alle blevet en adgangsbillet til det russiske folks miljø.
Denne tilgang, rejst til flaget af adskillige Russophobic a la "menneskerettighedsorganisationer" og de russiske Russophobic Massemedier, har oversvømmet æteren. Men Putin og andre som ham vil før eller siden stadig være nødt til at stå til ansvar for deres ord om ydmygelse af det russiske folk. Forskernes dom er nådesløs:
1) I 2009 blev en komplet "læsning" (sekventering) af genomet af en repræsentant for den russiske etnos afsluttet. Det vil sige, at sekvensen af alle seks milliarder nukleotider i den russiske mands genom er blevet bestemt. Hele hans genetiske økonomi er nu i fuld udsyn.
(Det menneskelige genom består af 23 par kromosomer: 23 - fra moderen, 23 - fra faderen. Hvert kromosom indeholder ét DNA-molekyle dannet af en kæde på 50-250 millioner nukleotider. Genomet fra en russisk mand blev sekventeret. Center "Kurchatov Institute", på initiativ af korresponderende medlem af det russiske videnskabsakademi, direktør for det nationale forskningscenter "Kurchatov Institute" Mikhail Kovalchuk Ifølge oplysninger modtaget på det russiske videnskabsakademi, kun til køb af udstyr til sekventering, Kurchatov Institute brugte omkring $ 20 millioner. Center "Kurchatov Institute" har en anerkendt videnskabelig status i verden.)
Det er kendt, at dette er det syvende afkodede genom bag Ural-ryggen: før det var der yakuter, buryater, kinesere, kasakhere, gamle troende, Khanty. Det vil sige, at alle forudsætningerne for det første etniske kort over Rusland er blevet skabt. Men alle disse var så at sige sammensatte genomer: stykker samlet efter afkodning af det genetiske materiale fra forskellige repræsentanter for den samme befolkning.
Det komplette genetiske portræt af en bestemt russisk mand er kun det ottende i verden. Nu er der nogen at sammenligne russere med: en amerikaner, en afrikaner, en koreaner, en europæisk ...
« Vi fandt ingen mærkbare tatariske bidrag i russisk genom, hvilket afviser teorier om den destruktive indflydelse af det mongolske åg, - understreger lederen af den genomiske retning ved National Research Center "Kurchatov Institute", akademiker Konstantin Skryabin. -Sibirere er genetisk identiske med gamle troende, de har ét russisk genom. Der er ingen forskelle mellem russernes og ukrainernes genomer - ét genom... Vores forskelle med polakkerne er ringe."
Akademiker Konstantin Skryabin mener, at "et genetisk kort over alle folkeslag i verden vil blive udarbejdet om fem til seks år - dette er et afgørende skridt i retning af at forstå enhver etnisk gruppes modtagelighed for stoffer, sygdomme og fødevarer." Føl hvad det koster ... Amerikanerne i 1990'erne gav følgende skøn: omkostningerne ved at sekventere et nukleotid - $ 1; ifølge andre kilder - op til $ 3-5.
(Sekvenseringen (stavningen af den genetiske kode) af mitokondrie-DNA og DNA'et fra det menneskelige Y-kromosom er den mest avancerede DNA-analysemetode til dato. "Stået af træet i Østafrika. Og Y-kromosomet er kun hos mænd og overføres derfor praktisk talt uændret til mandlige afkom, mens alle andre kromosomer, når de overføres fra far og mor til deres børn, blandes af naturen, som et sæt kort, før de bliver uddelt. , i modsætning til indirekte tegn (udseende, kropsproportioner), sekventering af mitokondrie-DNA og DNA fra Y-kromosomet er indiskutabel og direkte indikativ for graden af menneskers slægtskab.)
2) Fremragende antropolog, forsker af menneskets biologiske natur, A.P. Bogdanov skrev i slutningen af det 19. århundrede: "Vi bruger ofte udtryk: dette er rent russisk skønhed, dette er en spyttende hare, et typisk russisk ansigt. Man kan være overbevist om, at ikke noget fantastisk, men virkeligt ligger i dette generelle udtryk for russisk fysiognomi. I hver af os, i vores "ubevidstes" sfære, er der et ret bestemt koncept af den russiske type "(AP Bogdanov" Antropologisk fysiognomi ". M., 1878).
Hundrede år senere, og nu kommer den moderne antropolog V. Deryabin, ved hjælp af den nyeste metode til matematisk multivariat analyse af blandede træk, til samme konklusion: "Den første og vigtigste konklusion er at fastslå russernes betydelige enhed. i hele Rusland og umuligheden af at identificere selv de tilsvarende regionale typer, klart afgrænset fra hinanden ”(“Spørgsmål om antropologi. ”Udgave 88, 1995). Hvordan kommer denne russiske antropologiske enhed til udtryk, enheden af arvelige genetiske egenskaber udtrykt i en persons udseende, i kroppens struktur?
Først og fremmest - hårfarve og øjenfarve, formen af strukturen af kraniet. Ifølge disse tegn adskiller vi os, russerne, sig fra både de europæiske folk og mongoloiderne. Og vi kan slet ikke sammenlignes med negrene og semitterne, uoverensstemmelserne er for slående. Akademiker V.P. Alekseev beviste en høj grad af lighed i kraniets struktur blandt alle repræsentanter for det moderne russiske folk, idet han samtidig specificerede, at den "proto-slaviske type" er meget stabil og går tilbage til den neolitiske æra og muligvis den mesolitiske . Ifølge antropologen Deryabins beregninger findes lyse øjne (grå, gråblå, blå og blå) hos russere hos 45 procent, i Vesteuropa kun 35 procent lyse øjne. Mørkt, sort hår hos russere findes hos fem procent, hos befolkningen i udenlandsk Europa - hos 45 procent. Den konventionelle visdom om russernes "snus-nosedness" er heller ikke bekræftet. Hos 75 procent af russerne findes en lige næseprofil.
Konklusion fra antropologiske videnskabsmænd:
"Russere er i deres racesammensætning typiske kaukasiere, der indtager en central position blandt Europas folk ved de fleste antropologiske karakteristika og adskiller sig i en lidt lysere pigmentering af øjne og hår. Det er også nødvendigt at anerkende den betydelige enhed af den racemæssige type russere i hele det europæiske Rusland."
"En russer er en europæer, men en europæer med fysiske egenskaber, der kun er særegne for ham. Disse tegn udgør, hvad vi kalder en typisk hare."
Antropologer har for alvor ridset russeren, og - der er ingen tatar, altså en mongoloid, i russerne. Et af de typiske tegn på en mongoloid er epicanthus - en mongolsk fold i den indre øjenkrog. Hos typiske mongoloider findes denne fold hos 95 procent; i undersøgelsen af otte og et halvt tusinde russere blev en sådan fold fundet hos kun 12 mennesker og i sin embryonale form.
Et andet eksempel. Russerne har bogstaveligt talt et særligt blod - overvægten af 1. og 2. gruppe, hvilket fremgår af den langsigtede praksis med blodtransfusionsstationer. Hos jøder, for eksempel, er den dominerende blodgruppe den 4., den negative Rh-faktor er mere almindelig. I biokemiske blodprøver viste det sig, at russere, som alle europæiske folk, har et specielt PH-c-gen, dette gen er praktisk talt fraværende hos mongoloider (OV Borisova "Polymorfisme af erytrocytsyrefosfatase i forskellige befolkningsgrupper i Sovjetunionen." "Spørgsmål om antropologi". Udgave 53, 1976).
Det viser sig, at uanset hvordan du ridser en russer, stadig ikke en tatar, vil du ikke finde nogen anden i ham. Dette bekræftes af encyklopædien "Peoples of Russia", i kapitlet "The Racial Composition of the Population of Russia" bemærkes det: "Repræsentanter for den kaukasiske race udgør mere end 90 procent af landets befolkning og omkring 9 procent er repræsentanter for former blandet mellem kaukasiere og mongoloider. Antallet af rene mongoloider overstiger ikke 1 million mennesker." ("Folk i Rusland". M., 1994).
Det er let at beregne, at hvis der er 84 procent af russerne i Rusland, så er de alle udelukkende mennesker af den europæiske type. Folkene i Sibirien, Volga-regionen, Kaukasus og Ural er en blanding af de europæiske og mongolske racer. Dette var perfekt udtrykt af antropologen A.P. Bogdanov i det 19. århundrede, hvor han studerede folkene i Rusland, skrev han, og afviste fra sin langt, langt væk dagens myte om, at russerne hældte en andens blod ind i deres folk under invasioner og kolonisering:
"Måske giftede mange russere sig med indfødte kvinder og blev bosat, men de fleste af de primitive russiske kolonialister i hele Rusland og Sibirien var ikke sådan. De var et købmand, industrifolk, der brød sig om at indrette sig på deres egen måde, i overensstemmelse med deres eget velstandsideal. Og dette ideal om den russiske mand er slet ikke sådan, at det nemt kan fordreje sit liv med en slags "skrald", da den russiske mand nu alt for ofte ærer hedningen. Han vil gøre forretninger med ham, vil være kærlig og venlig med ham, vil blive glad for ham i alt, bortset fra at blive beslægtet, for at introducere et fremmed element i hans familie. Simple russiske mennesker er stadig stærke til dette, og når det kommer til familien, til etableringen af hans hjem, her har han en slags aristokrati. Ofte bor bosættere af forskellige stammer i nabolaget, men ægteskaber mellem dem er sjældne."
Gennem årtusinder har den russiske fysiske type forblevet stabil og uændret og har aldrig været en krydsning mellem forskellige stammer, der til tider beboede vores land. Myten er blevet aflivet, vi må forstå, at blodkaldet ikke er en tom sætning, at vores nationale idé om den russiske type er den russiske races virkelighed. Vi skal lære at se denne race, beundre den, værdsætte den i vores nære og fjerne russiske slægtninge. Og så, måske, vores russiske appel til helt fremmede, men vores eget folk for os - far, mor, bror, søster, søn og datter, vil blive genoplivet. Vi er jo faktisk alle fra en enkelt rod, fra én slags - den russiske slags.
3) Antropologer var i stand til at identificere udseendet af en typisk russisk person. For at gøre dette var de nødt til at overføre alle fotografier fra fotobiblioteket på Museum of Anthropology til en enkelt skala med fuld-ansigts- og profilbilleder af typiske repræsentanter for befolkningen i de russiske regioner i landet og kombinere dem i henhold til øjnenes pupiller, overlejret på hinanden. De endelige fotografier blev selvfølgelig slørede, men gav en idé om udseendet af det russiske referencefolk. Dette var den første virkelig sensationelle opdagelse. Lignende forsøg fra franske videnskabsmænd førte trods alt til et resultat, som de var nødt til at skjule for borgerne i deres land: efter tusindvis af kombinationer af de opnåede fotografier af reference Jacques og Marianne, så de på grå ansigtsløse ovaler af ansigter. Et sådant billede kunne, selv blandt franskmændene, der er fjernest fra antropologien, rejse et unødvendigt spørgsmål: findes der overhovedet en fransk nation?
Desværre gik antropologer ikke længere end at skabe fotografiske portrætter af typiske repræsentanter for den russiske befolkning i forskellige regioner i landet og overlejrede dem ikke på hinanden for at opnå udseendet af en absolut russisk mand. Til sidst blev de tvunget til at indrømme, at de kunne få problemer på arbejdet for sådan et billede. Forresten blev de "regionale" skitser af det russiske folk kun offentliggjort i den almindelige presse i 2002, og før det blev de kun offentliggjort i små udgaver i videnskabelige publikationer for specialister. Nu kan du selv vurdere, hvor ligner de de typiske filmiske Ivanushka og Marya.
Desværre tillader for det meste sort-hvide gamle arkivfotos af det russiske folks ansigter ikke at formidle det russiske folks højde, fysik, farve på huden, håret og øjnene. Men antropologer har skabt et verbalt portræt af russiske mænd og kvinder. De er af medium bygning og medium højde, lysebrunhårede med lyse øjne - grå eller blå. Forresten, i løbet af forskningen blev der også opnået et verbalt portræt af en typisk ukrainer. Standardukraineren adskiller sig kun fra russeren i farven på hans hud, hår og øjne - han er en mørkhudet brunette med regelmæssige ansigtstræk og brune øjne. En snub næse viste sig at være absolut ukarakteristisk for den østslaviske (findes kun hos 7% af russere og ukrainere), dette tegn er mere typisk for tyskere (25%).
4) I 2000 tildelte den russiske fond for grundforskning omkring en halv million rubler fra statsbudgetmidler til undersøgelse af det russiske folks genpulje. Det er umuligt at gennemføre et seriøst program med en sådan finansiering. Men det var mere et skelsættende end blot en økonomisk beslutning, hvilket indikerer en ændring i landets videnskabelige prioriteter. For første gang i russisk historie var forskere fra Laboratory of Human Population Genetics ved Medical Genetics Center i det russiske akademi for medicinske videnskaber, som modtog et tilskud fra den russiske fond for grundforskning, i stand til fuldt ud at fokusere på undersøgelsen af genpuljen af det russiske folk, og ikke små folk, i tre år. Og begrænset finansiering ansporede kun deres opfindsomhed. De supplerede deres molekylærgenetiske undersøgelser med en analyse af frekvensfordelingen af russiske efternavne i landet. Denne metode var meget billig, men dens informationsindhold oversteg alle forventninger: en sammenligning af geografien af efternavne med geografien af genetiske DNA-markører viste deres næsten fuldstændige sammenfald.
Desværre kunne fortolkningerne af familieanalysen, der dukkede op i medierne efter den første offentliggørelse af dataene i et specialiseret videnskabeligt tidsskrift, skabe et forkert indtryk af målene og resultaterne af videnskabsmænds enorme arbejde. Lederen af projektet, Doctor of Sciences Elena Balanovskaya, forklarede, at det vigtigste ikke var, at efternavnet Smirnov viste sig at være mere almindeligt blandt det russiske folk end Ivanov, men at der for første gang var en komplet liste over ægte russiske efternavne. udarbejdet for landets regioner. Først blev der udarbejdet lister for fem betingede regioner - nord, central, central-vest, central-øst og syd. I alt var der i alle regioner omkring 15 tusinde russiske efternavne, hvoraf de fleste kun blev fundet i en af regionerne og var fraværende i andre. Da regionale lister blev lagt oven på hinanden, identificerede videnskabsmænd kun 257 såkaldte "helrussiske efternavne". Interessant nok besluttede de på den sidste fase af undersøgelsen at tilføje efternavnene på indbyggerne i Krasnodar-territoriet til listen over den sydlige region, idet de forventede, at overvægten af ukrainske efternavne af efterkommerne af Zaporozhye-kosakkerne, smidt ud her af Catherine II , ville reducere den al-russiske liste markant. Men denne yderligere begrænsning reducerede listen over al-russiske efternavne med kun 7 enheder - til 250. Hvorfra den åbenlyse og ikke for alles behagelige konklusion fulgte, at Kuban hovedsageligt var beboet af russiske mennesker. Og hvor blev ukrainerne af, og var der overhovedet nogen ukrainere - et stort spørgsmål.
I tre år gik deltagerne i projektet "Russian Gene Pool" rundt med en sprøjte og et reagensglas næsten hele Den Russiske Føderations europæiske territorium og lavede en meget repræsentativ prøve af russisk blod.
Imidlertid var billige indirekte metoder til at studere det russiske folks genetik (ved efternavne og dermatoglyffer) kun hjælpemidler til den første undersøgelse af genpuljen af den titulære nationalitet i Rusland. Hans vigtigste molekylærgenetiske resultater er tilgængelige i monografien "Russian Gene Pool" (Publishing House "Luch"). Desværre måtte videnskabsmænd på grund af manglende offentlig finansiering udføre en del af undersøgelsen sammen med deres udenlandske kolleger, som indførte et moratorium for mange af resultaterne, indtil fælles publikationer blev offentliggjort i den videnskabelige presse. Intet forhindrer os i at beskrive disse data med ord. Så på Y-kromosomet er den genetiske afstand mellem russere og finner 30 konventionelle enheder. Og den genetiske afstand mellem en russisk person og de såkaldte finsk-ugriske folk (Mari, Veps osv.), der bor på Den Russiske Føderations territorium, er 2-3 enheder. Kort sagt er de genetisk næsten identiske. Resultaterne af analysen af mitokondrielt DNA viser, at russerne fra tatarerne er i samme genetiske afstand på 30 konventionelle enheder, som adskiller os fra finnerne, men mellem ukrainerne fra Lvov og tatarerne er den genetiske afstand kun 10 enheder .... Og samtidig er ukrainere fra venstrebredden Ukraine genetisk lige så tæt på russerne som komi-zyryanerne, mordoverne og Mari.
Har sprog gener? - Hvorfor har genpuljer brug for navne? - Hvad siger afstandskort? - Ikke ét kort, men en hel fan!§en. Afstande fra tre sprogfamilier: FRA INDO-EUROPÆISK: forskellene vokser mod øst - Men de fleste af bestandene er genetisk tætte; - FRA URAL: afstandene vokser fra øst til vest - Men mange befolkninger er tætte - finsk-ugrisk substrat blandt slaverne og tyrkerne; - FRA ALTAI: kun tæt på sig selv - Ingen indflydelse på naboer i Europa
§2. Afstande fra russere, hviderussere, ukrainere: Klassiske markører - Nordrussere er længere fra almindelige russere end ukrainere, mordovere og tjuvasjer - Autosomale DNA-markører - Forrige billede - Næsten alle er tæt på russere - Bortset fra Kaukasus og Ural - Y-kromosom - Samme billede med større kontrast - Afstande fra hviderussere - Kun tilsvarende for slaverne - Det samme billede for ukrainerne - Så det er for russerne, at de østeuropæiske befolkninger er tæt på, og ikke for slaverne generelt!
HAR SPROG GENER?
Vi vil gerne svare med det samme, at forfatterne ligesom læseren ved, at sprog ikke har gener. Dette er forståeligt selv på det daglige plan - hvor mange russere, spredt rundt i verden af bølger af den første, anden og anden emigration, taler en række forskellige sprog! Og de har alle de samme gener, arvet fra deres forfædre.
Hvorfor taler vi så om generne fra den slaviske eller germanske sprogfamilie? Er det videnskabeligt? Temmelig. Når alt kommer til alt, er vi engageret i befolkningsgenetik og taler kun om befolkningen af mennesker, der taler sprogene i den slaviske eller germanske gren af sprog. Og der er ikke andet bag de "sproglige navne".
Vi har allerede sagt mere end én gang, at befolkninger er flerlagede og kan være af meget forskellige rækker - fra elementære befolkninger (flere nabolandsbyer) til hele menneskehedens befolkning. Alle disse er populationer, og de er indlejret i hinanden som rededukker: mange populationer af lavere rang passer ind i populationen af den næste højere rang, og så videre. Vi definerer groft en af disse mellemliggende matryoshka-populationer efter etnicitet. Det er den eneste grund til, at vi kan tale om den russiske genpulje – altså om den befolkning, der er præget af menneskers tilhørsforhold til det russiske folk. Desuden bestemmes dette tilhørsforhold af folket selv, og på ingen måde af genetik! Og først efter at folk identificerede sig selv som russere eller som nordmænd (eller sagde, at deres bedsteforældre tænkte over det), begynder genetikere at se upartisk: er befolkningerne af russere og nordmænd forskellige, og hvor meget forskellige fra hinanden? Vi kalder betinget sådanne populationer "russiske" eller "norske", mens vi fuldt ud er klar over, at genpuljer og populationer er biologiske enheder, som vi giver "humanitære" navne.
Men lad os understrege, fordi vi har navnene "russisk" eller "norsk" for genpuljer af datoer, betyder det ikke, at "russiske gener" eller "norske gener" pludselig dukkede op på scenen! Der er ingen "russiske" eller "ukrainske" gener, ligesom der ikke er nogen slaviske eller romerske gener. Nej, om ikke andet fordi generne er meget ældre end menneskerne og er spredt ud over næsten hele verden. Disse spørgsmål diskuterer vi dog i bogens afslutning (kapitel 10). Og nu er det kun vigtigt for os at besvare spørgsmålet - hvis der ikke er nogen russiske eller slaviske gener, hvorfor kalder vi så genpuljer sådanne navne?
Hvorfor har genpuljer brug for navne?
Kun fordi populationer (og deres genpuljer) skal have tydelige navne. Du kan selvfølgelig lade genpuljen være unavngiven og hele tiden gentage "den største gamle befolkning på landet i regionerne i den østeuropæiske slette og mere nordlige regioner, groft korreleret med grænserne for den russiske stat før Ivan den Forfærdelige ." Men selv fra en sådan sætning vil det forblive uklart, hvem vi analyserer (for eksempel om vi inkluderer karelere, Izhora, tatarer eller mordovere). Og hvis vi siger (som vi beskrev i detaljer i begyndelsen af bogen), at vi med den russiske genpulje mener oprindelige russere på landet i deres "urgangs" (historiske) område, og derefter bruger udtrykket "russisk genpulje" i hele bog, så er det lettere for læseren at forstå, hvad forfatterne taler om. Derfor giver vi genpuljerne konventionelle navne - for at lette forståelsen.
Men for at give navne til matryoshka-dukker af en højere rang, er det nødvendigt at bruge en slags befolkningsklassificering. I kapitel 2 testede vi for eksempel, hvor genetisk effektive racemæssige og sproglige klassifikationer er. Og blandt folkene i Sibirien blev den genetiske effektivitet af klassificeringen af folk efter typen af ornament og efter typerne af shaman tamburiner kontrolleret. Og det viste sig, at ornamentet identificerer populationer dårligt, men shamantamburiner er ikke mindre effektive til at identificere befolkninger end sprog. Ikke desto mindre er den sproglige klassifikation blevet udviklet mest detaljeret. Derfor er navnene på befolkninger ofte givet ved navne på sprog. Så det er nu accepteret i de biologiske videnskaber. Og når vi for eksempel taler om det "finsk-ugriske" lag af genpuljen, forstår både antropologer og arkæologer os. De forstår, at vi taler om en bestemt befolkning af mennesker, ret udvidet i tid og rum. Og det gør ikke noget, at nu har chuvasherne ændret deres tidligere sprog til tyrkisk, og det gør ikke noget, at vi ikke ved, hvilket sprog de gamle befolkninger talte, hvis de ikke efterlod skriftlige beviser. En enorm række data fra forskellige videnskaber (herunder for eksempel toponymi - navnene på floder eller søer) vidner om, at der var et samfund af mennesker her, som vi nu giver det foreløbige navn på den "finsk-ugriske" verden.
Derfor, i både dette og næste afsnit, hvor vi sammenligner genetiske afstande fra populationer med "sproglige" navne, forråder vi ikke videnskaben, men følger dens strenge regler. Vi tager den sproglige klassifikation af folkeslag; så giver vi i overensstemmelse med den hver gruppe af populationer et konventionelt "sprogligt" navn; og endelig beregner vi de gennemsnitlige genfrekvenser for de populationer fra denne gruppe, der lever i det analyserede territorium. Og så ser vi på, hvordan hver enkelt befolkning i Østeuropa adskiller sig fra disse gennemsnitlige frekvenser af "indoeuropæiske" eller "altai"-populationer. Samtidig er forfatterne, ligesom læseren, klar over, at Altai-sprogene i Europa tales af folk med et helt andet fysisk udseende - fra Gagauz til Kalmyks. Men vi har ikke ret til på dette grundlag at udelukke nogen fra de grupper, som sprogvidenskaben har identificeret – vi opregner ærligt, hvilke folkeslag der indgår i befolkningen med et givet "sprogligt" navn.
HVAD FORTÆLLER DISTANCEKORT OM?
Genetiske afstandskort er næsten vigtigere end hovedkomponentkort. Disse to grundlæggende genogeografiske værktøjer, brugt sammen, giver en komplementær beskrivelse af genpuljen. Hovedkomponentkort giver os mulighed for at fremsætte hypoteser om de faktorer, der dannede de observerede mønstre, og kort over genetiske afstande giver os mulighed for at teste disse hypoteser.
Hvert kort over genetiske afstande givet i dette afsnit er gennemsnittet for alle undersøgte loci (tabel 8.1.1.). Det viser tydeligt, hvor genetisk tæt hver population i området er på én befolkningsgruppe specificeret af forskeren. En sådan gruppe af befolkninger kaldes "reference".
Genpuljen kan stilles spørgsmål: hvilke populationer er genetisk tæt på den befolkningsgruppe, der er af interesse for os? Hvilke er relativt afsidesliggende? Og hvilke er i øvrigt fundamentalt forskellige fra referencegruppen for hele sæt af genfrekvenser? Og kortet over genetiske afstande vil give svaret: hvor meget hvert punkt på kortet er genetisk tæt på eller langt fra referencegruppen. Vi vil se det med vores egne øjne.
Afstandskort adskiller sig fra den sædvanlige brug af genetiske afstande i kun én, men den vigtigste funktion: ved kortlægning inkluderer analysen befolkningens område, det vil sige det geografiske, rumlige aspekt.
Genetiske afstandskort afslører ofte forholdet mellem genetiske og geografiske afstande. Kortet demonstrerer, hvordan befolkningen i tilstødende og mere fjerntliggende territorier med afstand fra referencepopulationen (givet af forskeren) bliver genetisk mere og mere forskellig fra referencepopulationen. Denne stigning i genetisk afstand afhænger dog ikke kun af geografisk afstand. Ellers ville ethvert kort over genetiske afstande bestå af koncentriske cirkler, svarende til dem, der spredes på vand fra en kastet sten.
I virkeligheden kan afstande i én retning dog vokse hurtigt, hvilket indikerer barrierer for genflow; i andre retninger kan afstandene næppe øges, hvilket viser den genetiske affinitet af disse tilstødende grupper. I en række tilfælde kan det glatte forløb af isoliner blive forstyrret, og en genetisk fjern population identificeres blandt genetisk nære grupper, hvilket for eksempel kan indikere dens migration til dette territorium. At plotte genetiske afstande på et kort kan således give værdifuld information om forholdet mellem den undersøgte gruppe og resten af befolkningen i regionen, tilstedeværelsen af genetiske strømme, genetiske barrierer og relaterede grupper. Desuden modtager vi information om selve referencegruppen (for eksempel russere eller hviderussere): om genetisk diversitet inden for dens grænser, om afvigelser fra gennemsnitlige værdier inden for dens eget område.
IKKE ET KORT. EN HEL FAN!
Kortlægning af genetiske afstande tydeliggør mange træk ved genpuljen - især hvis vi ikke betragter et kort over afstande (fra ét folk), men en række kort - fra forskellige folkeslag, fra større befolkningsgrupper. Hvert nyt kort vil fortælle om den genetiske position af et nyt folk eller en gruppe af folk i den generelle genpulje i regionen. Sammenligning af hele fanen af kort vil vise, hvor stort bidraget fra hver af disse grupper er til den østeuropæiske genpulje, og hvor zonerne for deres blanding ligger.
Vi vil ikke her overveje kortene over genetiske afstande fra alle mennesker i Østeuropa - så vi ville gå for langt ud over omfanget af bogen om den russiske genpulje. Mere informative kort over afstande fra grupper af folk, relateret til hinanden. De afslører ikke mønstrene for individuelle folkeslags etnogenese, men de generelle begivenheder i dannelsen af befolkningen i Østeuropa. Som vi diskuterede i kapitel 2, er genogeografi baseret på princippet om "skalering": efterhånden som omfanget af de undersøgte grupper øges, afsløres spor af mere ældgamle og større begivenheder.
Derfor lægges der vægt på kort over afstande fra grupper af folkeslag. I §1 er kort konstrueret ud fra de gennemsnitlige frekvenser af gener hos folkene i de indoeuropæiske, uraliske og altaiiske sprogfamilier, der bor i Østeuropa. Derefter (§2) betragter vi kort over afstande fra det russiske folk, der viser dets position i den generelle genpulje i Østeuropa. Og afslutningsvis, lad os tage et kig på kortene over to andre folkeslag i Østeuropa - hviderussere og ukrainere, som historisk set er tæt på russiske befolkninger og kan have en lignende genpulje.
Alle kort læses ens. Jo mere et givet punkt på kortet er genetisk fjernet fra referencepopulationen, jo større afstanden er, jo mere intens er farven på dette punkt. Derfor er de letteste områder de områder med de mindste afstande. Det er de populationer, der ligner benchmark mest. De mørkeste er områderne med størst afstand. Det er populationer, der ikke genetisk ligner referencepopulationen. Selvfølgelig. så snart vi tager en anden referencepopulation, vil de samme punkter på kortet rapportere, at de har forskellige afstande til den nye reference. For at lette læsningen er alle afstandskort bygget i en enkelt skala, så du trygt kan sammenligne ikke kun forskellige dele af det samme kort, men også forskellige kort med hinanden efter farveintensitet.
§en. Afstande fra tre sprogfamilier
Overvej kortene over de genetiske afstande for alle befolkninger i Østeuropa fra folkene i de indoeuropæiske, uraliske og altaiiske sprogfamilier, der bor i det. For kortheds skyld præsenterer vi kort over ét "øjenvidne" - autosomale DNA-markører, da kortene over genetiske afstande med klassiske markører, som vi vil se i næste afsnit, er ret ens.
FRA FOLKET I DEN INDOEUROPÆISKE SPROGFAMILIE (DNA-MARKERE)
Et kort over genetiske afstande fra den indoeuropæiske sprogfamilie er vist i fig. 8.3.1.
Kortet blev bygget sådan her. For det første blev de gennemsnitlige frekvenser af DNA-markører beregnet for repræsentanter for den indoeuropæiske familie i Østeuropa: befolkninger af russere, ukrainere, hviderussere og moldovere. Derefter blev de gennemsnitlige "indoeuropæiske" genfrekvenser opnået på basis af dem. Yderligere beregnes de genetiske afstande fra disse gennemsnitlige "indoeuropæiske" frekvenser til frekvenserne på hvert punkt på kortet, og de opnåede afstandsværdier placeres i de samme noder på kortet.
Derfor, hvis værdierne for genetiske afstande for eksempel i det meste af Hviderusland, i distrikterne Kiev og Lvov falder i området fra 0,01 til 0,02 (fig. 8.3.1.), betyder det, at disse er ( i gennemsnit for alle gener) forskellene mellem disse populationer fra gennemsnitshyppighederne for folkene i den indoeuropæiske familie. Tværtimod er forskellene mellem Kalmyks, Komi og Bashkirs meget større - værdierne af genetiske afstande i deres bosættelses territorier er mere end 0,05 og 0,06. Resten af kortene over genetiske afstande læses på samme måde.
Kortet viser, som man kunne forvente, at befolkningen af russere i Centralrusland, ukrainere, hviderussere og moldovere (det vil sige selve de indoeuropæiske befolkninger) er tæt på gennemsnitsfrekvenserne for de indoeuropæiske folk i Østeuropa. . Men alt er ikke forudsigeligt - de nordrussiske befolkninger (selv om de også er indoeuropæere) adskiller sig markant fra de "gennemsnitlige indoeuropæere" - i samme grad som de ikke-indoeuropæiske folkeslag i den midterste Volga (Mari, Mordover, Chuvash) og det vestlige Kaukasus. Endelig er den mest fremragende befolkningen i Ural (især Komi), såvel som steppefolkene (bashkirerne, kalmykerne).
Lad os være opmærksomme på befolkningen i det russiske folk. De repræsenterer den indoeuropæiske sprogfamilie i Østeuropa, deres frekvenser blev brugt til at beregne de gennemsnitlige "indoeuropæiske" frekvenser. Og ikke desto mindre ser vi slående forskelle mellem russiske befolkninger i graden af nærhed til deres egen referencepopulation. Dette indikerer igen, at niveauet af heterogenitet af det russiske folks genpulje er så stort, at det tydeligt manifesteres selv på en østeuropæisk skala.
Generelt afsløres et klart geografisk mønster: når man bevæger sig mod øst, stiger afstandsværdierne gradvist, befolkningerne er mere og mere forskellige fra de gennemsnitlige karakteristika for de indoeuropæiske folk og folkene i den østlige udkant af Europa viser sig at være det mest genetisk ulige dem. Men generelt er de fleste af befolkningen i Østeuropa (inklusive Ural- og Kaukasiske befolkninger) tæt på de indoeuropæiske folk: den gennemsnitlige værdi af genetiske afstande på kortet er lille d = 0,028.
FRA FOLKET I URAL SPROG-FAMILIEN (DNA-MARKERE)
Det følgende kort over genetiske afstande er bygget ud fra de gennemsnitlige frekvenser af gener fra den uralske sprogfamilie og viser et andet billede (fig. 8.3.2.).
Fra Ural-familien er kun de østfinsktalende folkeslag (komier, udmurtere, mari, mordovere) blevet undersøgt med DNA-markører. Minimumsafstandene findes i disse folks bosættelses område, hovedsageligt i Ural. Tværtimod er befolkningen i den vestlige del af den russiske slette og Ciscaucasia genetisk fjernt fra de gennemsnitlige Ural-frekvenser. De midterste regioner i Østeuropa, der geografisk støder op til Ural, er tættere på Ural-folkene og genetisk.
Så, de mindste værdier af afstandene er lokaliseret i Ural og stiger gradvist længere mod vest. Sandsynligvis afspejler de områder, der er besat af mellemværdier, området for de gamle finsk-ugriske stammer assimileret af slaverne [Alekseeva, 1965]. Det er mærkeligt, at områderne for de tyrkisk-talende folk i Ural er tæt på karakteristikaene for den uraliske familie, hvilket forklares af den betydelige andel af det uraliske substrat i genpuljen af chuvasherne, tatarerne og nogle grupper af Bashkirs [Roginsky, Levin, 1978].
Den gennemsnitlige afstand på kortet, selv om den er højere end den "indoeuropæiske", er lille (d = 0,039). Dette bekræfter den betydelige repræsentation af den Ural-talende genpulje i den generelle østeuropæiske genpulje, som i vid udstrækning består af Ural-substratet.
FRA FOLKET I ALTAI LANGUAGE FAMILIE (DNA-MARKERE)
Det næste kort (Fig. 8.3.3.) Demonstrerer forskellene mellem hver østeuropæisk befolkning og folkene i Altai-sprogfamilien. Denne familie i Østeuropa er hovedsageligt repræsenteret af tyrkisktalende folk - kun kalmykere taler et sprog, der tilhører den mongolske gruppe af denne familie.
To tidligere kort over genetiske afstande (fra de indoeuropæiske og fra Ural-familierne) var karakteriseret ved små gennemsnitlige værdier af afstandene. På kortene (fig. 8.3.1., 8.3.2.) kunne dette mærkes ved overvægten af lyse toner. Tværtimod, på kortet over afstande fra Altai-familien (Fig. 8.3.3.), dominerer en mørk farve, svarende til den betydelige genetiske afstand for de fleste af de østeuropæiske populationer fra genpuljen i Altai-sprogfamilien. Kun områderne for folkene i Altai-sprogfamilien selv er naturligt tæt på deres gennemsnitlige værdier. Og umiddelbart uden for zonen for deres bosættelse viser resten af de østeuropæiske befolkninger sig at være skarpt forskellige fra de genetiske egenskaber hos de Altai-talende folk.
Dette afspejles i den større værdi af genetiske afstande end for tidligere kort. I gennemsnit på kortet var de d = 0,064, hvilket er næsten tre gange højere end den analoge værdi for indoeuropæiske folkeslag.
Således er indflydelsen fra folkene i Altai-familien på den østeuropæiske genpulje kun begrænset af zonen for deres bosættelse, og ifølge de undersøgte data spores praktisk talt ikke selv i tilstødende territorier. Denne kendsgerning kan forklares med den relativt sene optræden i Østeuropa af mange stammer, der taler Altai-familiens sprog [Nationer og religioner i verden, 1999], mens både indoeuropæiske og uraliske familier er sprogene i den ældre befolkning i Østeuropa [Cheboksarov, Cheboksarova, 1971; Bunak, 1980].
§2. Afstande fra russere, hviderussere, ukrainere
Så vi har lært den vigtigste "sammensætning" af den østeuropæiske genpulje - hvad er de vigtigste subgenofonds i den, i hvilke "andele" de er "blandet", og hvordan disse andele adskiller sig i forskellige dele af Østeuropa. Nu kan vi vende tilbage til hovedemnet i vores bog og overveje, hvordan alle østeuropæiske befolkningers stilling er i forhold til russerne? Da dette er et førende emne, vil vi give genetiske afstande fra russiske populationer for tre typer markører - klassiske markører, autosomale DNA-markører og markører for Y-kromosomet. Og for ikke at forveksle "rent russiske" funktioner med "slaviske", vil vi også overveje kort over afstande fra historisk tætte østslaviske folk - hviderussere og ukrainere.
FRA RUSSISK BEFOLKNING (KLASSISKE MARKØRER)
Kortet over genetiske afstande fra de gennemsnitlige russiske frekvenser ved klassiske markører viser graden af lighed mellem hver population i Østeuropa med den russiske genpulje. Det lyse område af den nærmeste nærhed til de centrale russiske genfrekvenser optager mellembåndet i Østeuropa - fra Hviderusland til den midterste Volga (fig. 8.3.4.). Mørke toner er områder, der er genetisk fjernt fra russerne. Der er relativt få af dem - i rækkefølge efter graden af afstand fra centralrusserne - det er Krim- og Sortehavsområdet, Nedre Volga, de baltiske stater, det russiske nord, Fennoskandia og de genetisk fjerne Ural.
Hviderussernes og ukrainernes levesteder viser ligheder med den russiske genpulje. De skarpe genetiske forskelle mellem det russiske nord og det nordøstlige Europa generelt, inklusive Vyatka, en gammel Novgorod-koloni, er overraskende.
Naturligvis bærer den oprindelige russiske befolkning, der bor i disse territorier, nu i videst udstrækning træk af en assimileret befolkning. Det er dog utroligt, at bidraget fra den finsk-ugriske befolkning her var højere end i populationerne af mordoverne og chuvasherne, som på kortet var fuldstændig inkluderet i den "centralrussiske genetiske region". Der er tre mulige kilder til sådanne forskelle. For det første kan selve det finsk-ugriske substrat trække mod de vestfinsktalende folk og ikke mod de østlige.
For det andet, som de arkæologiske data indikerer [Sedov, 1999], havde den novgorodiske kolonisering en anden oprindelse for de slaviske stammer selv. Dette betyder, at ikke kun substratet, men også det slaviske superstratum kan være unikt i det russiske nord. For det tredje, i de små nordlige populationer, er gendriftsfaktoren kraftigere, hvilket også kunne "transportere" dem væk fra det russiske hovedkontinent. Mest sandsynligt har alle tre faktorer virket parallelt, men opgaven med fremtidig forskning er at finde ud af deres virkelige forhold. Her kan homogene markører være til stor hjælp, der er med til at differentiere migrationsstrømme i rum og tid.
Nærheden til de "Centralrussiske" frekvenser manifesteres af de mest forskellige dele af det russiske område, inklusive dem, hvor de modsatte yderpunkter af hovedkomponenterne i den østeuropæiske genpulje er placeret (afsnit 8.2.). Et lignende billede kan forklares ud fra hypotesen om, at de "centralrussiske" frekvenser i sig selv i det væsentlige er "Centraleuropæiske", og den russiske genpulje er dannet af blandinger af de mest forskelligartede østeuropæiske komponenter (finsk-ugriske, slaviske, Østersøen osv.). Denne hypotese bekræftes også i kortene over genetiske afstande fra ukrainere, hviderussere og russere ifølge den mest informative DNA-markør - haplogrupper af Y-kromosomet.
FRA DEN RUSSISKE BEFOLKNING (AUTOSOMISKE DNA-MARKERE)
Som ifølge dataene om klassiske markører (fig. 8.3.4) er befolkningen i det centrale Rusland igen tæt på de centralrussiske genfrekvenser (fig. 8.3.5.). Hviderussere, der praktisk talt ikke kan skelnes fra de gennemsnitlige russiske karakteristika med hensyn til frekvenserne af klassiske markører, og ifølge DNA-data viser små forskelle. Befolkningen i Ural, Kaukasus, Volga-regionen og i mindre grad det russiske nord er meget forskellige fra de gennemsnitlige russiske frekvenser. I alle hovedpunkterne fører brugen af DNA og klassiske markører således til lignende resultater. Forskellene mellem de to kort skyldes efter vores mening hovedsageligt graden af kendskab til markører af forskellige typer, og det kan forventes, at efterhånden som data om DNA-polymorfi akkumuleres, vil billedet af deres variabilitet i stigende grad nærme sig de opnåede resultater vha. klassiske markører.
Den gennemsnitlige genetiske afstand for de østeuropæiske populationer fra de russiske frekvenser er lille (d = 0,28), hvilket kan være resultatet af en langvarig interaktion mellem den russiske genpulje og miljøet. Husk på, at afstanden fra indoeuropæiske folk generelt er karakteriseret ved den samme gennemsnitsværdi (d = 0,28). Når man sammenligner disse kort (Fig. 8.3.1. Og 8.3.5), bliver deres betydelige lighed indlysende. Dette er forståeligt, da russerne også er indo-europæere, og frekvenserne i de russiske befolkninger var inkluderet i beregningen for de indoeuropæiske folk. Det er mærkeligt, at forskellene mellem de russiske befolkninger mellem Volga og Vyatka, markeret på kortet over afstande fra middelfrekvenserne for indoeuropæiske folk, også er bevaret på kortet over afstande fra de gennemsnitlige russiske frekvenser.
Så den russiske genpulje viser sig at være tæt forbundet med genpuljen fra mange østeuropæiske folkeslag - med hensyn til genfrekvenser er hviderussiske, ukrainske, mordoviske og mange andre østeuropæiske befolkninger ekstremt tæt på russerne. Først når vi nærmer os Kaukasus og Ural, bliver populationens genpulje klart anderledes end gennemsnitskarakteristikaene for den russiske genpulje. Dette resultat er ikke uventet, da den russiske bosættelse over store territorier og den intensive udveksling af gener uden for det "oprindelige" område med de omkringliggende folk er indlysende. Det virker snarere interessant, at tilstedeværelsen af to bjergbarrierer (Kaukasus og Ural) optrådte på de gen-geografiske kort, hvilket til en vis grad begrænsede denne rumlige udvidelse af genpuljen.
FRA RUSSISK BEFOLKNING (Y-KROMOSOMMARKERE)
Dette kort adskiller sig på to måder. For det første ser vi hele Europa på det og ikke kun dets østlige halvdel (kortet er bygget på basis af de otte kort over individuelle haplogrupper, der blev overvejet i afsnit 6.3). For det andet er Y-kromosommarkørernes differentieringsevne meget højere, så forskellene mellem russiske befolkninger og naboer er mere udtalte. På trods af selv en "bredere" skala af intervaller dominerer intervallet af maksimale afstande på kortet - ifølge Y-kromosommarkører er næsten hele Europa væsentligt forskelligt fra den russiske genpulje (fig. 8.3.6). Kun de russiske befolkninger og hviderusserne selv er tættest på de gennemsnitlige russiske frekvenser, ukrainerne, vestslaviske folk (polakker, tjekkere, slovakker) og folkene i Volga-regionen viser en gennemsnitlig grad af nærhed. Som i de tidligere kort viser de nordrussiske populationer et udtalt særpræg, der adskiller sig skarpt fra den gennemsnitlige russiske genpulje.
Vi ser, at markørerne for Y-kromosomet bekræfter de tidligere identificerede mønstre af lighed mellem den "centralrussiske" genpulje med andre østslaviske folk og folkene i Volga-regionen og forskellene i det russiske nord. Y-kromosomets høje informationsindhold gør disse mønstre mere konvekse end for andre typer markører, og overvejelser på en skala af hele Europa tilføjer listen over folkeslag, der ligner den russiske genpulje af folk og polakker.
FRA BELARUSSER (KLASSISKE MARKØRER)
På tidligere kort (fig. 8.3.4., 8.3.5., 8.3.6.) så vi, at mange populationer i Østeuropa ligner den russiske genpulje.
Det er vigtigt at forstå: er alle disse populationer tæt på den russiske genpulje eller på en bred vifte af østslaviske populationer? Med andre ord: er hemmeligheden bag denne lighed i det russiske folks etniske historie eller i udvidelsen af de østlige slaver som helhed, og muligvis i den "indledende", før udvidelsen, ligheden mellem det slaviske og finsk-ugriske gen pools?
For at besvare dette spørgsmål analyserede vi den østeuropæiske genpools nærhed til hviderusserne - en anden østslavisk etnisk gruppe, meget tæt på det russiske folk i geografi, etnogenese og antropologisk type.
I fig. 8.3.7. et kort over de genetiske afstande for befolkninger i Østeuropa fra de gennemsnitlige hviderussiske genfrekvenser for et stort sæt klassiske genmarkører - 57 alleler af 21 loci er præsenteret. Vi ser et klart billede, fundamentalt forskelligt fra arten af variationen i den russiske genpulje. Næsten alle regioner, hvis befolkning viser den tætteste nærhed til den hviderussiske genpulje, er placeret på Hvideruslands territorium. Uden for det hviderussiske område stiger genetiske afstande hurtigt til betydelige værdier, hvilket indikerer klare genetiske forskelle mellem genpuljen af hviderussere og den østeuropæiske genpulje som helhed.
Kortet fanger den genetiske originalitet af genpuljen af hviderussere, hvilket indikerer den høje følsomhed af den genetiske afstandsmetode. Det skal bemærkes, at klare forskelle mellem den hviderussiske genpulje og genpuljen i naboterritorier er et vigtigt uventet resultat, eftersom antropologiske data normalt ikke afslører udtalte forskelle mellem hviderussere og nabogrupper [Alekseeva, 1973; Deryabin, 1999]. Selvfølgelig er denne genetiske ejendommelighed ved hviderussere meget relativ: den manifesterer sig kun i hviderussisk skala, som gennem et mikroskop, takket være den enorme opløsning af kort for at se selv fine detaljer. Husk på, at hviderussere i en anden skala - på kortene over genetiske afstande fra russere - praktisk talt ikke kan skelnes fra russere i det centrale Rusland. Under alle omstændigheder ligner hviderusserne meget mere dem end de russiske befolkninger i det russiske nord selv.
I modsætning til den russiske er den hviderussiske genpulje således ikke tæt på den østeuropæiske genpulje som helhed. Følgelig er den høje genetiske lighed mellem russiske befolkninger og befolkningen i de fleste østeuropæiske territorier ikke et træk, der er fælles for alle østslaviske folk, men dets egen karakteristik af den russiske genpulje.
FRA HVIDERUSSER (MARKERE OG KROMOSOMER)
Denne konklusion bekræftes af dataene om Y-kromosomet. Afstandskortet fra hviderussere (fig. 8.3.8.) er bygget i samme skala af intervaller som fra russere (fig. 8.3.6.). Men zonen, der genetisk ligner den hviderussiske genpulje, er mærkbart mindre: den omfatter kun slaviske folk (både østslaver, bortset fra det vestlige Ukraine og vestslaviske befolkninger), men inkluderer ikke folkene i Volga- og Ural-regionerne. Genetisk fællestræk med ikke-slavisk-talende befolkninger i Østeuropa er således den russiske genpuljes "privilegium" i modsætning til genpuljen af hviderussere, som adskiller sig markant fra disse folkeslag i Volga-regionen og Ural.
FRA UKRAINIERE (MARKERE OG KROMOSOMER)
For fuldstændigheden af overvejelserne om de østslaviske folk præsenterer vi også et kort over afstande fra ukrainerne (fig. 8.3.9.). Det ligner meget det kort, der lige er blevet gennemgået fra hviderusserne, kun zonen med maksimal nærhed er flyttet til området for ukrainerne selv, og denne zone inkluderer også den sydlige russiske og hviderussiske befolkning. Og de ikke-slaviske folkeslag i Østeuropa, som er relativt tæt på de russiske befolkninger, er lige så langt fra den ukrainske genpulje, som de er fra hviderussernes genpulje. Dette bekræfter rigtigheden af vores fortolkning, at den slaviske kolonisering af den østeuropæiske slette, ledsaget af assimileringen af den finsk-ugriske befolkning, primært involverede forfædrene til den moderne russiske befolkning fra hele det slaviske massiv.






Forskere har for første gang lavet et detaljeret kort over den genetiske struktur af britiske befolkninger. Det viste sig, at befolkningen i det keltiske territorium slet ikke er genetisk homogen, og den ældste genpulje er bevaret i Wales. En sammenligning af DNA fra britiske og kontinentale europæere pegede på den genetiske profil af forfædrene for hver region. I den moderne befolknings genpulje var det muligt at se afspejlingen af de vigtigste begivenheder i historien om bosættelsen på de britiske øer.
At studere de genetiske forskelle mellem moderne befolkninger hjælper med at se dybt ind i historien og se sporene af migration af befolkningsgrupper, takket være hvilken den moderne genpulje blev dannet. Dette arbejde blev udført af en international gruppe af forskere ledet af britiske eksperter fra University of Oxford og University College London, deres artikel blev offentliggjort i tidsskriftet Nature den 19. marts. Walter Bodmer, Peter Donnelly og deres kolleger analyserede i detaljer den genetiske sammensætning af den britiske befolkning. De byggede ikke kun et meget nøjagtigt genetisk kort over den moderne britiske befolkning, men fandt også spor af de vigtigste historiske migrationer og tydeliggjorde karakteren af bosættelsen på de britiske øer.
Genpuljen afspejler geografi
Forskerne indsamlede DNA fra 2.039 mennesker fra forskellige dele af Storbritannien. Når de valgte, blev de styret af følgende regel: alle menneskelige forfædre i tredje generation (både bedstemødre og begge bedstefædre) måtte ikke bo længere end 80 km fra hinanden. Denne tilstand gjorde det muligt at få et "øjebliksbillede" af den britiske befolknings genetiske struktur i slutningen af det 19. og begyndelsen af det 20. århundrede.
For at sammenligne DNA brugte forskerne mere end 500.000 enkeltnukleotidpolymorfier (SNP'er) spredt ud over genomet. Deres analyse blev udført ved hjælp af de statistiske metoder fineSTRUCTURE og GLOBETROTTER. Disse metoder kategoriserede DNA-prøver i klynger baseret på deres lighed, uanset hvor de blev indsamlet i Storbritannien. Og ved at placere en prøve af hver person på et punkt midtvejs fra deres bedsteforældres fødesteder, var de i stand til at se, hvordan genetisk lighed korrelerer med geografisk placering.
Genetiske klynger, der grupperede 2039 DNA-prøver fra britiske befolkninger.
Tilfældighederne var slående. Som det kan ses på figuren, var 17 genetiske klynger fordelt på kortet i overensstemmelse med den geografiske placering af de udtagne prøver. Forskellige klynger overlapper praktisk talt ikke hinanden. Og da disse klynger blev dannet fra alle prøver uden at tage hensyn til deres geografi, betyder det, at denne korrespondance afspejler befolkningens reelle genetiske mangfoldighed, understreger forfatterne.
Befolkningen i amtet Orkney på Orkneyøerne nord for Skotland viste sig at være den mest genetisk mangfoldige; den omfatter tre klynger. Genetiske forskelle er tydelige på tværs af Storbritannien: Nordengland, Skotland og Nordirland adskiller sig kollektivt fra det sydlige England, Nordwales adskiller sig fra Sydwales, og Cornwall danner en særskilt klynge. Klyngegrænser falder ofte sammen med de moderne grænser for historiske provinser, såsom Cornwall-Devon-grænsen, England-Wales-grænsen. Den største klynge (angivet med røde firkanter på kortet) dækker det meste af det centrale og sydlige England og østkysten. Det kombinerer næsten halvdelen af DNA-prøverne inkluderet i undersøgelsen.
Forfatterne byggede et fylogenetisk træ af britiske befolkninger, der viste, hvordan klynger blev dannet, hvordan hovedgrenene blev adskilt. Som du kan se i træet, splittede en gren af befolkningen på Orkneyøerne sig på det tidligste stadie, og i næste fase splittede Wales sig ud. Efterfølgende gafler i træet førte til adskillelsen af Nord- og Sydwales, adskillelsen af det nordlige England, Skotland og Nordirland fra resten af England og adskillelsen af Cornwall fra resten af klyngerne.
Således blev ideen om den genetiske homogenitet af de keltiske befolkninger i Storbritannien som den ældste, ikke påvirket af den saksiske migration, ikke bekræftet. Tværtimod viste den keltiske del af Storbritannien (Skotland, Nordirland, Wales og Cornwall) sig at være meget mere genetisk forskelligartet end resten af territoriet.
Selvom klyngerne i den britiske befolkning skilte sig så tydeligt ud, da forskerne målte den genetiske afstand mellem dem, viste den sig at være lille (gennemsnit 0,002, maksimum 0,007). Det betyder, at de observerede forskelle er meget subtile og kun kan ses med den stærke "forstørrelse", som forskerne har skabt ved hjælp af en kraftfuld statistisk metode. Til sammenligning analyserede de deres data ved hjælp af standardmetoder - principal component analysis (PCA) og ADMIXTURE. Begge metoder viste adskillelse af grene fra Orkney og Wales, men afslørede ikke den finere genetiske struktur af befolkningen.
Kontinentalt fodaftryk i det britiske genom
Genetiske forskelle inden for den britiske befolkning kan være resultatet af isoleringen af visse grupper af befolkningen fra hinanden og afspejler desuden bidraget fra forskellige migrationer og processen med at blande migranter med den oprindelige befolkning. For at håndtere sidstnævnte brugte forskerne den samme metode til at sammenligne DNA-prøver fra den britiske befolkning med prøver fra mere end 6.000 kontinentale europæere fra 10 lande. Ifølge genetisk lighed blev disse prøver opdelt i 51 grupper.

Europæiske genetiske profiler for 17 britiske klynger.
For hver britisk klynge vurderede videnskabsmænd bidraget fra hver kontinental gruppe og bestemte forfædrenes genetiske profil. Analysen viste, at nogle europæiske grupper, såsom dem fra det vestlige Tyskland, det flamske Belgien, det nordvestlige Frankrig, det sydlige Frankrig og Spanien, bidrog væsentligt til oprindelsen af stort set alle britiske klynger. Andre har kun bidraget til nogle specifikke klynger, for eksempel havde nordmanden en betydelig indflydelse på dannelsen af klynger i Orkney og mindre i Skotland og Nordirland.
Genpuljen afspejler historien
Genetiske data har afspejlet spor af migrationer til de britiske øer i løbet af de sidste 10 tusind år. Ifølge arten af indflydelsen fra kontinentale grupper på britiske klynger skelner forskerne det relative tidspunkt for migrationsbegivenheder. De grupper, hvis bidrag findes i alle klynger, indikerer tydeligvis mere gamle migrationer. Der er gået nok tid siden da til, at deres haplotyper spredte sig til andre dele af Storbritannien. Det kontinentale fodaftryk i individuelle klynger ser ud til at være resultatet af nyere migrationer.

De vigtigste begivenheder i bosættelsen af de britiske øer.
Det mest bemærkelsesværdige mærke blev efterladt af angelsaksernes erobring af Storbritannien i det 5. århundrede. Det genetiske bidrag fra angelsaksisk migration er betydeligt i det sydøstlige, centrale og sydlige England, selvom det ikke overstiger 50% nogen steder, og i det meste af territoriet varierer det fra 10 til 40%. Dette indikerer, at angelsakserne ikke fuldstændigt fortrængte den lokale befolkning af briterne (kelterne), men blandede sig med den. Adskillige klynger falder topografisk sammen med de angelsaksiske kongerigers og klaners territorier. Det ser ud til, at disse territorier har bevaret deres etnografiske og genetiske identitet i mange århundreder, understreger forskerne.
I befolkningerne på Orkneyøerne tilskrives 25% af genpuljen forfædrene af norsk oprindelse, disse er spor efter de norske vikingers invasion af øerne, som skete i det 9. århundrede. Genetiske data indikerer også, at de nordiske vikinger ikke fortrængte den oprindelige befolkning, men blandede sig med den. Men det var ikke muligt at finde klare genetiske spor efter de danske vikinger, som samtidig kontrollerede en væsentlig del af England.
Hvad angår den tidligste bosættelse af de britiske øer - efter afslutningen af den sidste istid - er dens spor bedst bevaret i befolkningen i Wales. Dataene peger også på en efterfølgende betydelig migration fra kontinentet, selv før Romerrigets erobring af Storbritannien i begyndelsen af det første årtusinde e.Kr. Den genetiske indflydelse af denne migration spredte sig over hele England, Skotland og Nordirland, men omgik praktisk talt Wales, som har bevaret den ældste genpulje.
Undersøgelsens forfattere har endegyldigt vist, hvordan genetik kan supplere og forfine arkæologisk og sproglig information for at rekonstruere befolkningernes historie. For at få et detaljeret genetisk kort over landet og se ind i den britiske befolknings fortid, blev de hjulpet af kompetent udvælgelse af indledende data og kraftfulde statistiske metoder.
En kilde:
Den britiske befolknings genetiske struktur i fin skala
Stephen Leslie, Bruce Winney, Garrett Hellenthal, Dan Davison, Abdelhamid Boumertit, Tammy Day, Katarzyna Hutnik, Ellen C. Royrvik, Barry Cunliffe, Wellcome Trust Case Control Consortium, International Multiple Sclerosis Genetics Consortium, Daniel J. Lawson, Daniel Falush, Colin Freeman, Matti Pirinen, Simon Myers, Mark Robinson, Peter Donnelly & Walter Bodmer
Nature, 2015, v.519, 7543, doi: 10.1038 / nature14230
http://www.nature.com/nature/journal/v519/n7543/full/nature14230.html









