Solusykli. Lyhyesti välivaiheista
Ihmisen kehon korkeus johtuu solujen koon ja lukumäärän kasvusta, jälkimmäisen takaa jakautumisprosessi tai mitoosi. Solujen lisääntyminen tapahtuu solunulkoisten kasvutekijöiden vaikutuksesta, ja solut itse käyvät läpi toistuvan tapahtumasarjan, joka tunnetaan solusyklinä.
Niitä on neljä pääasiallista vaiheet: G1 (esynteettinen), S (synteettinen), G2 (postsynteettinen) ja M (mitoottinen). Tätä seuraa sytoplasman ja plasmakalvon erottaminen, mikä johtaa kahteen identtiseen tytärsoluun. Vaiheet Gl, S ja G2 ovat osa välivaihetta. Kromosomireplikaatio tapahtuu synteettisen vaiheen tai S-vaiheen aikana.
Suurin osa soluja eivät ole alttiina aktiiviselle jakautumiselle, niiden mitoottinen aktiivisuus vaimenee GO-vaiheen aikana, joka on osa G1-vaihetta.
M-vaiheen kesto on 30-60 minuuttia, kun taas koko solusykli tapahtuu noin 20 tunnissa Iästä riippuen normaalit (ei-kasvain) ihmissolut käyvät läpi jopa 80 mitoottista sykliä.
Prosessit solusykli Niitä kontrolloidaan peräkkäin toistuvalla avainentsyymeillä, joita kutsutaan sykliinistä riippuvaisiksi proteiinikinaaseiksi (CDPK:iksi), sekä niiden kofaktoreilla, sykliineillä. Tässä tapauksessa fosfokinaasien ja fosfataasien vaikutuksesta tapahtuu erityisten sykliini-CZK-kompleksien fosforylaatiota ja defosforylaatiota, jotka ovat vastuussa syklin tiettyjen vaiheiden alkamisesta.
Lisäksi asiaankuuluvalla samanlaisia vaiheita kuin CZK-proteiinit aiheuttavat kromosomien tiivistymistä, ydinvaipan repeytymistä ja solun tukirangan mikrotubulusten uudelleenorganisoitumista fissiokaran (mitoottisen karan) muodostamiseksi.
solusyklin G1-vaihe
G1-vaihe- M- ja S-vaiheiden välivaihe, jonka aikana sytoplasman määrä kasvaa. Lisäksi G1-vaiheen lopussa on ensimmäinen tarkistuspiste, jossa tarkistetaan DNA:n korjaus- ja ympäristöolosuhteet (ovatko ne riittävän suotuisat S-vaiheeseen siirtymiselle).
Jos kyseessä on ydinvoima DNA vaurioituneena p53-proteiinin aktiivisuus lisääntyy, mikä stimuloi p21:n transkriptiota. Jälkimmäinen sitoutuu spesifiseen sykliini-CZK-kompleksiin, joka on vastuussa solun siirtämisestä S-faasiin, ja estää sen jakautumista Gl-vaiheessa. Tämä sallii korjausentsyymien korjata vaurioituneita DNA-fragmentteja.
Jos patologioita esiintyy Viallisen DNA:n p53-proteiinin replikaatio jatkuu, mikä sallii jakautuvien solujen keräävän mutaatioita ja edistää kasvainprosessien kehittymistä. Tästä syystä p53-proteiinia kutsutaan usein "genomin vartijaksi".
solusyklin G0-vaihe
Solujen lisääntyminen nisäkkäissä on mahdollista vain muiden solujen erittämien solujen osallistuessa. solunulkoiset kasvutekijät, jotka vaikuttavat proto-onkogeenien kaskadisignaalitransduktion kautta. Jos solu ei G1-vaiheen aikana vastaanota asianmukaisia signaaleja, se poistuu solusyklistä ja siirtyy G0-tilaan, jossa se voi olla useita vuosia.
G0-esto tapahtuu proteiinien avulla - mitoosin vaimentajat, joista yksi on retinoblastoomaproteiini(Rb-proteiini), jota koodaavat retinoblastoomageenin normaalit alleelit. Tämä proteiini kiinnittyy vinoon sääteleviin proteiineihin ja estää solujen lisääntymiseen tarvittavien geenien transkription stimulaation.
Solunulkoiset kasvutekijät tuhoavat eston aktivoitumalla Gl-spesifiset sykliini-CZK-kompleksit, jotka fosforyloivat Rb-proteiinin ja muuttavat sen konformaatiota, minkä seurauksena yhteys säätelyproteiineihin katkeaa. Samaan aikaan viimeksi mainitut aktivoivat koodaamiensa geenien transkription, mikä laukaisee proliferaatioprosessin.
solusyklin S-vaihe
Vakiomäärä DNA-kaksoisheliksit jokaisessa solussa vastaava yksijuosteisten kromosomien diploidijoukko on yleensä merkitty 2C:ksi. 2C-sarja säilyy koko G1-vaiheen ajan ja kaksinkertaistuu (4C) S-vaiheen aikana, kun uutta kromosomaalista DNA:ta syntetisoidaan.
Alkaen lopusta S-vaihe ja M-vaiheeseen asti (mukaan lukien G2-faasi), jokainen näkyvä kromosomi sisältää kaksi tiukasti sitoutunutta DNA-molekyyliä, joita kutsutaan sisarkromatideiksi. Siten ihmissoluissa on S-vaiheen lopusta M-vaiheen puoliväliin 23 paria kromosomeja (46 näkyvää yksikköä), mutta 4C (92) tuman DNA:n kaksoiskierrettä.
Käynnissä mitoosi identtiset kromosomisarjat jakautuvat kahden tytärsolun kesken siten, että kukin niistä sisältää 23 paria 2C-DNA-molekyylejä. On huomattava, että G1- ja G0-faasit ovat ainoat solusyklin vaiheet, joiden aikana 46 kromosomia soluissa vastaa 2C-sarjaa DNA-molekyylejä.
solusyklin G2-vaihe
Toinen tarkista kohta, jossa solukokoa testataan, on G2-vaiheen lopussa, S-vaiheen ja mitoosin välissä. Lisäksi tässä vaiheessa, ennen kuin siirrytään mitoosiin, replikaation täydellisyys ja DNA:n eheys tarkistetaan. Mitoosi (M-vaihe)
1. Prophase. Kromosomit, joista kukin koostuu kahdesta identtisestä kromatidista, alkavat tiivistyä ja tulevat näkyviksi ytimen sisällä. Solun vastakkaisissa napoissa karamainen laite alkaa muodostua tubuliinikuiduista kahden sentrosomin ympärille.
2. Prometafaasi. Ydinkalvo jakautuu. Kinetokorit muodostuvat kromosomien sentromeerien ympärille. Tubuliinikuidut tunkeutuvat ytimeen ja keskittyvät lähelle kinetokoreja yhdistäen ne senrosomeista lähteviin kuituihin.
3. Metafaasi. Kuitujen jännitys saa kromosomit asettumaan karan napojen puoliväliin, jolloin muodostuu metafaasilevy.
4. Anafaasi. Sentromeeri-DNA, joka on jaettu sisarkromatidien kesken, monistuu, ja kromatidit erottuvat ja siirtyvät lähemmäs napoja.
5. Telofaasi. Erotetut sisarkromatidit (jotka tästä eteenpäin katsotaan kromosomeiksi) saavuttavat navat. Jokaisen ryhmän ympärille ilmestyy ydinkalvo. Tiivistynyt kromatiini haihtuu ja muodostuu nukleolia.
6. Sytokineesi. Solukalvo supistuu ja keskelle napojen väliin muodostuu halkeamisuurre, joka ajan myötä erottaa kaksi tytärsolua.
Centrosomisykli
Sisään G1 vaiheaika jokaiseen sentrosomiin liitetty sentriolipari erottuu. S- ja G2-vaiheiden aikana muodostuu uusi tytärsentrioli vanhojen sentriolien oikealle puolelle. M-vaiheen alussa sentrosomi jakautuu ja kaksi tytärsentrosomia liikkuu kohti solunapoja.
Solusykli on solun olemassaolon ajanjakso sen muodostumishetkestä emosolun jakautuessa sen omaan jakautumiseen tai kuolemaan asti.
Solusyklin kesto
Solusyklin pituus vaihtelee eri solujen välillä. Nopeasti lisääntyvät aikuisten organismien solut, kuten orvaskeden ja ohutsuolen hematopoieettiset tai tyvisolut, voivat päästä solukiertoon 12-36 tunnin välein Lyhyitä solusyklejä (noin 30 minuuttia) havaitaan piikkinahkaisten ja sammakkoeläinten munien nopean pirstoutumisen aikana. ja muita eläimiä. Koeolosuhteissa monilla soluviljelylinjoilla on lyhyt solusykli (noin 20 tuntia). Useimmilla aktiivisesti jakautuvilla soluilla mitoosien välinen aika on noin 10-24 tuntia.
Solusyklin vaiheet
Eukaryoottisolusykli koostuu kahdesta jaksosta:
Solujen kasvujakso, jota kutsutaan "interfaasiksi", jonka aikana syntetisoidaan DNA:ta ja proteiineja ja valmistaudutaan solun jakautumiseen.
Solunjakautumisen jakso, jota kutsutaan "vaiheeksi M" (sanasta mitoosi - mitoosi).
Välivaihe koostuu useista jaksoista:
G 1-vaihe (englanniksi. aukko- aikaväli) tai alkukasvuvaihe, jonka aikana tapahtuu mRNA:n, proteiinien ja muiden solukomponenttien synteesi;
S-vaihe (englanniksi. synteesi- synteesi), jonka aikana tapahtuu soluytimen DNA-replikaatio, myös sentriolien kaksinkertaistuminen tapahtuu (jos niitä tietysti on).
G 2 -vaihe, jonka aikana tapahtuu valmistautuminen mitoosiin.
Erilaistuneissa soluissa, jotka eivät enää jakautu, solusyklissä ei välttämättä ole G1-vaihetta. Tällaiset solut ovat lepovaiheessa G0.
Solunjakautumisjakso (vaihe M) sisältää kaksi vaihetta:
karyokineesi (solutuman jakautuminen);
sytokineesi (sytoplasman jakautuminen).
Mitoosi puolestaan jaetaan viiteen vaiheeseen.
Solunjakautumisen kuvaus perustuu valomikroskopiatietoihin yhdistettynä mikrokuvaukseen sekä kiinnittyneiden ja värjäytyneiden solujen valo- ja elektronimikroskopian tuloksiin.
Solusyklin säätely
Säännöllinen muutossekvenssi solusyklin jaksoissa tapahtuu proteiinien, kuten sykliinistä riippuvaisten kinaasien ja sykliinien, vuorovaikutuksen kautta. G0-vaiheen solut voivat päästä solukiertoon, kun ne altistuvat kasvutekijöille. Erilaiset kasvutekijät, kuten verihiutaleperäiset, epidermaaliset ja hermokasvutekijät, sitoutumalla reseptoreihinsa laukaisevat solunsisäisen signalointikaskadin, mikä lopulta johtaa sykliinigeenien ja sykliinistä riippuvaisten kinaasien transkriptioon. Sykliinistä riippuvaiset kinaasit aktivoituvat vain ollessaan vuorovaikutuksessa vastaavien sykliinien kanssa. Erilaisten sykliinien pitoisuus solussa muuttuu koko solusyklin ajan. Sykliini on sykliini-sykliiniriippuvaisen kinaasikompleksin säätelykomponentti. Kinaasi on tämän kompleksin katalyyttinen komponentti. Kinaasit eivät ole aktiivisia ilman sykliinejä. Erilaisia sykliinejä syntetisoidaan solusyklin eri vaiheissa. Siten sykliini B:n pitoisuus sammakon munasoluissa saavuttaa maksiminsa mitoosin aikaan, kun sykliini B/sykliiniriippuvaisen kinaasikompleksin katalysoima fosforylaatioreaktioiden koko sarja käynnistyy. Mitoosin loppuun mennessä proteinaasit tuhoavat sykliinin nopeasti.
InterfaasiG1 seuraa mitoosin telofaasia. Tämän vaiheen aikana solu syntetisoi RNA:ta ja proteiineja. Vaiheen kesto on useista tunnista useisiin päiviin. G0. Solut voivat poistua syklistä ja olla G0-vaiheessa. G0-vaiheessa solut alkavat erilaistua. S. S-vaiheen aikana proteiinisynteesi jatkuu solussa, DNA:n replikaatio tapahtuu ja sentriolit erottuvat. Useimmissa soluissa S-vaihe kestää 8-12 tuntia. G2. G2-vaiheen aikana RNA- ja proteiinisynteesi jatkuu (esimerkiksi tubuliinin synteesi mitoottisia karan mikrotubuluksia varten). Tytärsentriolit saavuttavat lopullisten organellien koon. Tämä vaihe kestää 2-4 tuntia. Mitoosi Mitoosin aikana ydin (karyokineesi) ja sytoplasma (sytokineesi) jakautuvat. Mitoosin vaiheet: profaasi, prometafaasi, metafaasi, anafaasi, telofaasi (kuvat 2-52). Prophase. Jokainen kromosomi koostuu kahdesta sisarkromatidista, jotka on yhdistetty sentromeerillä, nukleoli katoaa. Sentriolit järjestävät mitoottisen karan. Sentriolipari on osa mi-
1. Mikä on solukierto?
Solusykli on solun olemassaolo sen muodostumishetkestä emosolun jakautumisen aikana sen omaan jakautumiseen (mukaan lukien tämä jakautuminen) tai kuolemaan. Solusykli koostuu interfaasista ja mitoosista (solun jakautumisesta).
2. Mitä kutsutaan välivaiheeksi? Mitä päätapahtumia esiintyy välivaiheen G 1 -, S- ja G 2 -jaksoissa?
Interfaasi on solusyklin osa kahden peräkkäisen jakautumisen välillä. Koko interfaasin aikana kromosomit ovat spiraalittomia ja sijaitsevat solun ytimessä kromatiinin muodossa. Pääsääntöisesti välivaihe koostuu kolmesta jaksosta:
● Esisynteettinen jakso (G 1) – välivaiheen pisin osa (2-3 tunnista useisiin päiviin). Tänä aikana solu kasvaa, organellien määrä lisääntyy, energiaa ja aineita kertyy DNA:n myöhempää kaksinkertaistamista varten. G 1 -jakson aikana jokainen kromosomi koostuu yhdestä kromatidista. Diploidisen solun kromosomien (n) ja kromatidien (c) joukko G1-jaksolla on 2n2c.
● Synteesijakson (S) aikana tapahtuu DNA:n kaksinkertaistamista (replikaatiota) sekä proteiinien synteesiä, jotka ovat välttämättömiä myöhempään kromosomien muodostumiseen. Tänä samana ajanjaksona sentriolit kaksinkertaistuvat. S-jakson loppuun mennessä jokainen kromosomi koostuu kahdesta identtisestä sisarkromatidista, jotka on yhdistetty sentromeeriin. Diploidisen solun kromosomi- ja kromatidijoukko S-jakson lopussa (eli replikaation jälkeen) on 2n4c.
● Postsynteettisen jakson (G 2) aikana solu kerää energiaa ja syntetisoi proteiineja tulevaa jakautumista varten (esimerkiksi tubuliinia rakentamaan mikrotubuluksia, jotka myöhemmin muodostavat karan). Koko G 2 -jakson ajan solun kromosomi- ja kromatidijoukko on 2n4c.
Interfaasin lopussa alkaa solujen jakautuminen.
3. Mitä soluja luonnehtii G 0 -jakso? Mitä tänä aikana tapahtuu?
Toisin kuin jatkuvasti jakautuvat solut (esimerkiksi ihon orvaskeden itukerroksen solut, punainen luuydin, eläinten maha-suolikanavan limakalvo, kasvien koulutuskudoksen solut), useimmat monisoluisen organismin solut ottavat erikoistumispolun ja G 1 -jakson osan läpäisyn jälkeen lepojakson aikana (G 0 -jakso).
G0-jakson solut suorittavat tietyt tehtävänsä kehossa, mutta niissä esiintyy aineenvaihdunta- ja energiaprosesseja, mutta valmistautuminen replikaatioon ei tapahdu. Tällaiset solut menettävät yleensä pysyvästi kykynsä jakautua. Esimerkkejä ovat neuronit, linssisolut ja monet muut.
Jotkut G0-jaksossa olevat solut (esimerkiksi leukosyytit, maksasolut) voivat kuitenkin poistua siitä ja jatkaa solusykliä käyden läpi kaikki interfaasi- ja mitoosijaksot. Siten maksasolut voivat jälleen saada jakautumiskyvyn useiden kuukausien lepojakson jälkeen.
4. Miten DNA:n replikaatio suoritetaan?
Replikaatio on DNA:n kaksinkertaistuminen, yksi templaattisynteesin reaktioista. Replikaation aikana erityiset entsyymit erottavat alkuperäisen emo-DNA-molekyylin kaksi juostetta ja rikkovat vetysidokset komplementaaristen nukleotidien välillä. DNA-polymeraasin, pääreplikaatioentsyymin, molekyylit sitoutuvat erotettuihin juosteisiin. Sitten DNA-polymeraasimolekyylit alkavat liikkua emoketjuja pitkin käyttämällä niitä templaatteina ja syntetisoivat uusia tytärketjuja, jotka valitsevat niille nukleotideja komplementaarisuuden periaatteen mukaisesti.
Replikaation tuloksena muodostuu kaksi identtistä kaksijuosteista DNA-molekyyliä. Jokainen niistä sisältää yhden alkuperäisen emomolekyylin ketjun ja yhden vasta syntetisoidun tytärketjun.
5. Ovatko homologiset kromosomit muodostavat DNA-molekyylit samoja? Sisarkromatidien koostumuksessa? Miksi?
Yhden kromosomin sisarkromatideissa olevat DNA-molekyylit ovat identtisiä (niillä on sama nukleotidisekvenssi), koska ne muodostuvat alkuperäisen DNA-emomolekyylin replikaation seurauksena. Kukin kahdesta DNA-molekyylistä, jotka muodostavat sisarkromatidit, sisältää yhden alkuperäisen DNA-emämolekyylin (templaatin) juosteen ja yhden uuden, tälle templaatille syntetisoidun tytärjuosteen.
DNA-molekyylit homologisissa kromosomeissa eivät ole identtisiä. Tämä johtuu siitä, että homologisilla kromosomeilla on eri alkuperä. Jokaisessa homologisessa kromosomiparissa toinen on äidin (peritty äidiltä) ja toinen isän (peritty isältä).
6. Mikä on nekroosi? Apoptoosi? Mitä yhtäläisyyksiä ja eroja nekroosin ja apoptoosin välillä on?
Nekroosi on solujen ja kudosten kuolema elävässä organismissa, joka johtuu erilaisten haitallisten tekijöiden vaikutuksesta.
Apoptoosi on kehon säätelemää ohjelmoitua solukuolemaa (niin sanottu "soluitsemurha").
Samankaltaisuudet:
● Nekroosi ja apoptoosi ovat kahdenlaisia solukuolemia.
● Esiintyy kaikissa kehon elämänvaiheissa.
Erot:
● Nekroosi on satunnainen (sopimaton) solukuolema, joka voi johtua altistumisesta korkeille ja matalille lämpötiloille, ionisoivalle säteilylle, erilaisille kemikaaleille (myös myrkkyille), mekaanisista vaurioista, heikentyneestä verenkierrosta tai kudosten hermotuksesta tai allergisesta reaktiosta. Keho suunnittelee alun perin apoptoosin (geneettisesti ohjelmoitu) ja säätelee sitä. Apoptoosin aikana solut kuolevat ilman suoria vaurioita, koska ne vastaanottavat tietyn molekyylisignaalin - "käskyn tuhota itsensä".
● Apoptoosin seurauksena yksittäiset tietyt solut kuolevat (vain ne, jotka ovat saaneet "määräyksen"), ja yleensä kokonaisia soluryhmiä kuolee nekroottisesti.
● Vaurioituneiden solujen nekroottisen kuoleman aikana kalvon läpäisevyys häiriintyy, proteiinisynteesi pysähtyy, muut aineenvaihduntaprosessit pysähtyvät, ydin, organellit ja lopulta koko solu tuhoutuvat. Tyypillisesti leukosyytit hyökkäävät kuoleviin soluihin, ja nekroosialueelle kehittyy tulehdusreaktio. Apoptoosin aikana solu hajoaa erillisiksi fragmenteiksi, joita ympäröi plasmalemma. Tyypillisesti valkosolut tai naapurisolut absorboivat kuolleiden solujen fragmentteja aiheuttamatta tulehdusreaktiota.
Ja (tai) muita tärkeitä ominaisuuksia.
7. Mikä on ohjelmoidun solukuoleman merkitys monisoluisten organismien elämässä?
Yksi apoptoosin päätehtävistä monisoluisessa organismissa on varmistaa solujen homeostaasi. Apoptoosin ansiosta erityyppisten solujen lukumäärän oikea suhde säilyy, kudosten uusiutuminen varmistetaan ja geneettisesti vialliset solut poistetaan. Apoptoosi näyttää keskeyttävän solujen jakautumisen äärettömän. Apoptoosin heikkeneminen johtaa usein pahanlaatuisten kasvainten ja autoimmuunisairauksien kehittymiseen (patologiset prosessit, joissa kehittyy immuunireaktio elimistön omia soluja ja kudoksia vastaan).
8. Miksi luulet, että suurimmassa osassa eläviä organismeja tärkein perinnöllisen tiedon säilyttäjä on DNA ja RNA suorittaa vain aputoimintoja?
DNA-molekyylin kaksijuosteinen luonne on sen itsensä monistumisen (replikaation) ja vaurioiden eliminoinnin - korjauksen prosessien taustalla (vaurioitumaton juoste toimii matriisina vaurioituneen juosteen palauttamiseksi). Koska RNA on yksijuosteinen, se ei kykene replikoitumaan, ja sen korjausprosessit estyvät. Lisäksi ylimääräisen hydroksyyliryhmän läsnäolo riboosissa (verrattuna deoksiriboosiin) tekee RNA:sta herkemmän hydrolyysille kuin DNA.
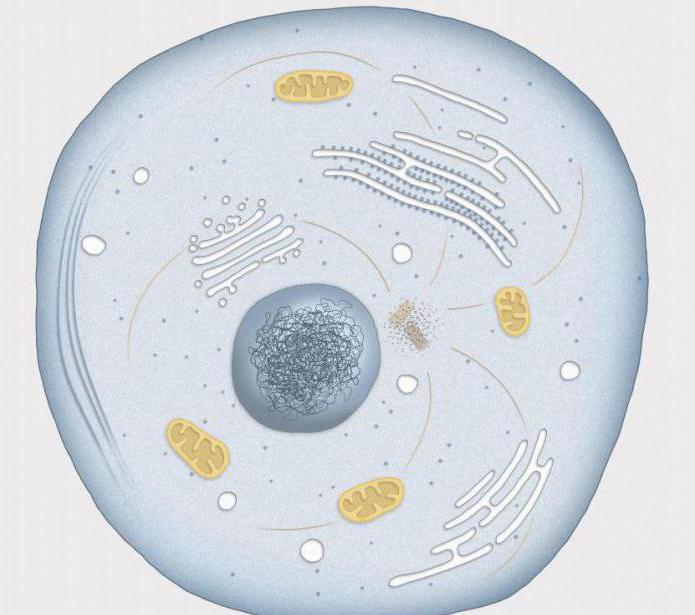
Mikä on interfaasi? Termi tulee latinan sanasta "inter", käännettynä "välillä", ja kreikan sanasta "phasis" - jakso. Tämä on tärkein ajanjakso, jonka aikana solu kasvaa ja kerää ravintoaineita valmistautuakseen seuraavaan jakautumiseen. Interfaasi vie suuren osan koko solusyklistä, jopa 90% solun koko elämästä.
Mikä on välivaihe
Pääsääntöisesti suurin osa solukomponenteista kasvaa koko vaiheen ajan, joten siinä on melko vaikeaa erottaa yksittäisiä vaiheita. Siitä huolimatta biologit ovat jakaneet interfaasin kolmeen osaan keskittyen replikaation aikaan solun ytimessä.
Vaihejaksot: G(1)-vaihe, S-vaihe, G(2)-vaihe. Synteettinen jakso (G1), jonka nimi tulee englanninkielisestä aukosta, käännettynä "väliksi", alkaa välittömästi jaon jälkeen. Tämä on erittäin pitkä ajanjakso, joka kestää kymmenestä tunnista useisiin päiviin. Juuri tänä aikana tapahtuu aineiden kertymistä ja valmistautuminen geneettisen materiaalin kaksinkertaistamiseen: RNA-synteesi alkaa ja tarvittavat proteiinit muodostuvat.
Mikä on välivaihe viimeisellä jaksollaan? Presynteettisessä vaiheessa ribosomien lukumäärä kasvaa, karkean endoplasmisen retikulumin pinta-ala kasvaa ja uusia mitokondrioita ilmaantuu. Solu, joka kuluttaa paljon energiaa, kasvaa nopeasti.
Erilaistuneet solut, jotka eivät enää pysty jakautumaan, pysyvät lepovaiheessa, jota kutsutaan G0:ksi.

Välivaiheen pääjakso
Riippumatta siitä, mitä prosesseja tapahtuu solussa interfaasin aikana, jokainen osavaihe on tärkeä mitoosin yleisen valmistelun kannalta. Synteesijaksoa voidaan kuitenkin kutsua käännekohtaksi, koska sen aikana kromosomit kaksinkertaistuvat ja alkaa välittömästi valmistautuminen jakautumiseen. RNA:n syntetisointia jatketaan, mutta se yhdistyy välittömästi kromosomiproteiinien kanssa, mikä aloittaa DNA:n replikaation.
Solun välivaihe tässä osassa kestää kuudesta kymmeneen tuntia. Tämän seurauksena jokainen kromosomi kaksinkertaistuu ja koostuu jo sisarkromatidiparista, jotka sitten hajoavat karan napoille. Synteettisessä faasissa sentriolit kaksinkertaistuvat, jos niitä tietysti on solussa. Tänä aikana kromosomit voidaan nähdä mikroskoopilla.

Kolmas jakso
Geneettisesti kromatidit ovat täysin identtisiä, koska yksi niistä on äidin ja toinen replikoituu käyttämällä lähetti-RNA:ta.
Heti kun kaiken geneettisen materiaalin täydellinen kaksinkertaistuminen on tapahtunut, alkaa synteesijälkeinen ajanjakso, joka edeltää jakautumista. Tämän jälkeen muodostuu mikrotubuluksia, joista myöhemmin muodostuu kara ja kromatidit hajaantuvat napoihin. Energiaa myös varastoituu, koska mitoosin aikana ravinteiden synteesi vähenee. Synteesijakson kesto on lyhyt, yleensä vain muutaman tunnin.

Tarkistuspisteet
Tämän prosessin aikana solun on läpäistävä tietyt tarkistuspisteet - tärkeät "merkit", jonka jälkeen se siirtyy toiseen vaiheeseen. Jos solu ei jostain syystä päässyt läpi tarkistuspisteen läpi, koko solusykli jäätyy, ja seuraava vaihe ei ala ennen kuin on korjattu ongelmat, jotka estivät sen kulkemisen tarkastuspisteen läpi.
Pääkohtia on neljä, joista suurin osa on vain välivaiheessa. Solu läpäisee ensimmäisen tarkastuspisteen esisynteettisessä vaiheessa, kun DNA:n eheys tarkistetaan. Jos kaikki on oikein, synteettinen jakso alkaa. Siinä sovintokohta on DNA:n replikaation tarkkuuden varmistaminen. Synteesin jälkeisen vaiheen tarkistuspiste on vaurioiden tai puutteiden tarkistus kahdessa edellisessä kohdassa. Tämä vaihe tarkistaa myös kuinka täydellinen replikaatio ja solut ovat tapahtuneet. Ne, jotka eivät läpäise tätä testiä, eivät saa osallistua mitoosiin.
Ongelmia välivaiheessa
Normaalin solusyklin häiriintyminen voi johtaa mitoosin epäonnistumiseen, mutta myös kiinteiden kasvainten muodostumiseen. Lisäksi tämä on yksi tärkeimmistä syistä niiden ulkonäköön. Kunkin vaiheen normaali kulku, olipa se kuinka lyhyt tahansa, ennalta määrää seuraavien vaiheiden onnistuneen loppuunsaattamisen ja ongelmien puuttumisen. Kasvainsoluissa on muutoksia solusyklin tarkistuspisteissä.
Esimerkiksi solussa, jossa on vaurioitunut DNA, interfaasin synteettistä jaksoa ei tapahdu. Tapahtuu mutaatioita, jotka johtavat p53-proteiinigeenien menetykseen tai muutoksiin. Soluissa ei ole solusyklin tukkeutumista, ja mitoosi alkaa etuajassa. Tällaisten ongelmien seurauksena on suuri määrä mutanttisoluja, joista useimmat ovat elämättömiä. Kuitenkin ne, jotka voivat toimia, synnyttävät pahanlaatuisia soluja, jotka voivat jakautua hyvin nopeasti lyhentyneen tai puuttuvan lepovaiheen vuoksi. Interfaasin ominaisuus sallii mutanttisoluista koostuvien pahanlaatuisten kasvainten jakautumisen niin nopeasti.
Vaiheiden välinen kesto
Otetaan muutama esimerkki siitä, kuinka paljon pidempään vaiheiden välinen jakso kestää solun elämässä verrattuna mitoosiin. Tavallisten hiirten ohutsuolen epiteelissä "lepovaihe" kestää vähintään kaksitoista tuntia, ja itse mitoosi kestää 30 minuutista tuntiin. Fabapavun juuren muodostavat solut jakautuvat 25 tunnin välein, ja M-vaihe (mitoosi) kestää noin puoli tuntia.
Mikä on solujen elämän välivaihe? Tämä on tärkein ajanjakso, jota ilman ei vain mitoosi, vaan myös solujen elämä kokonaisuudessaan olisi mahdotonta.









